
Cancer Heterogeneity and Plasticity ISSN 2818-7792
Cancer Heterogeneity and Plasticity 2024;1(2):0008 | https://doi.org/10.47248/chp2401020008
Original Research Open Access
Transcription factor AP-4 reactivates telomerase reverse transcriptase independently of genome alteration in non-HBV associated hepatocellular carcinoma
Huiju Lim
1,2,†
 ,
Trevor Kwan-Hung Wu
1,2,†
,
Trevor Kwan-Hung Wu
1,2,†
 ,
Eliana Mary Senires Suarez
1,2
,
Tiffany Ching-Yun Yu
1,2
,
Joyce Man-Fong Lee
1,2
,
Yu-Man Tsui
1,2
,
Daniel Wai-Hung Ho
1,2
,
Karen Man-Fong Sze
1,2
,
Irene Oi-Lin Ng
1,2
,
Eliana Mary Senires Suarez
1,2
,
Tiffany Ching-Yun Yu
1,2
,
Joyce Man-Fong Lee
1,2
,
Yu-Man Tsui
1,2
,
Daniel Wai-Hung Ho
1,2
,
Karen Man-Fong Sze
1,2
,
Irene Oi-Lin Ng
1,2
Correspondence: Karen Man-Fong Sze; Irene Oi-Lin Ng
Academic Editor(s): Dean G. Tang
Received: Sep 6, 2024 | Accepted: Nov 21, 2024 | Published: Nov 26, 2024
© 2024 by the author(s). This is an Open Access article distributed under the terms of the Creative Commons License Attribution 4.0 International (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium or format, provided the original work is correctly credited.
Cite this article: Lim H, Wu T, Suarez E, Yu T, Lee J, Tsui Y, Ho D, Sze K, Ng I. Transcription factor AP-4 reactivates telomerase reverse transcriptase independently of genome alteration in non-HBV associated hepatocellular carcinoma. Cancer Heterog Plast. 2024;1(2):0008. https://doi.org/10.47248/chp2401020008
Recurrent genome alterations have been identified in the telomerase reverse transcriptase (TERT) promoter region, a common occurrence in various types of cancer. In addition to epigenetic alterations at the TERT promoter region, the recruitment of transcription factors to this region, which potentially drives the reactivation of TERT gene transcription in human hepatocellular carcinoma (HCC) remains unknown. In this study, using in-silico DNA binding sequence analysis at the TERT promoter and with confirmation with dual luciferase reporter and chromatin immunoprecipitation assays, we found that Transcription Factor AP-4 (TFAP4) physically interacted with the TERT promoter, driving telomerase reactivation. Furthermore, TFAP4 mRNA is upregulated in human HCCs and positively correlates with TERT mRNA expression, according to TCGA-LIHC database, our in-house clinical samples, and HCC cell lines. Interestingly, the knockdown of TFAP4 only suppressed TERT expression levels and telomere length in HCC cells which are not associated with HBV infection. Additionally, we identified that the CCCTC-binding factor (CTCF) physically interacted with the TFAP4 promoter, leading to induced TFAP4 gene transcription in HCC. Clinically, TFAP4 mRNA expression significantly correlated with TERT mRNA expression in alcohol-related HCCs, but not in HBV-associated HCCs, according to TCGA-LIHC cohort. Consistently, ethanol enhanced both TFAP4 and TERT mRNA expression in non-HBV-associated HCC cells but not HBV-associated HCC cells. In conclusion, our findings demonstrate that TFAP4 directly regulates TERT gene transcription via CTCF in non-HBV-associated HCCs. However, its role in regulating telomerase expression or activity through HBV DNA integration in HBV-associated cells might be limited.
KeywordsTERT, TFAP4, CTCF, HCC
According to GLOBOCAN 2020, liver cancer is the third leading cause of death worldwide, with a high prevalence in East Asia [1]. Hepatocellular carcinoma (HCC) constitutes 75–85% of primary liver cancer in adults. The common risk factors of HCC include chronic hepatitis B virus (HBV) and hepatitis C virus (HCV) infection, chronic alcohol consumption, and metabolic disorders, and HBV and HCV infections together account for 80–90% of HCC cases [2]. Telomeres, which cap the ends of chromosomes, protect them from shortening after cell division. The enzyme telomerase reverse transcriptase (TERT) helps cancer cells maintain telomere length, thereby ensuring genomic integrity and the stability of chromosome structure for infinite cell division. Although TERT has been studied in cancer biology for decades, the mechanisms are not clearly characterized [3, 4].
Recent studies suggest that telomere length is associated with tumor progression and aggressiveness in human HCC [5]. Hypomethylation at the TERT promoter has been identified in human cell lines, including HCC cell lines, but not in human HCC [6]. Regarding transcription factor-mediated TERT gene transcription, MYC is known as a major regulator of TERT at the transcriptional level. It can directly modulate TERT expression during carcinogenesis by binding to the TERT promoter or cooperating with other transcription factors such as SP1 and ETS transcription factor family [7, 8]. Apart from the MYC transcription factor, NFKB1, JUN, STAT3, STAT5, PAX, and HIF1s are suggested to be transcriptional activators of the TERT promoter [6]. Moreover, several putative binding sites of transcription factors, including Transcription Factor AP-4 (TFAP4), have been identified on the TERT promoter, but no evidence suggests its role in regulating TERT gene transcription in HCC [9].
HBV DNA integration and promoter mutations at the TERT promoter leading to TERT mRNA expression have been extensively studied in HCC [10–14]. We previously showed that HBV-TERT promoter integration harnesses host transcription factor E74 like ETS transcription factor 4 (ELF4) in driving TERT mRNA expression in HBV-associated HCC [10]. However, other mechanisms leading to TERT reactivation in HCC are largely unknown. In the present study, we used in-silico DNA binding sequence analysis at the TERT promoter, coupled with dual luciferase reporter assay and chromatin immunoprecipitation assay, and discovered TFAP4 a novel transcription activator of the TERT promoter in HCC. Furthermore, we demonstrated that TFAP4 mRNA is upregulated and positively correlated with TERT mRNA expression in alcohol-related HCCs in The Cancer Genome Atlas Liver Hepatocellular Carcinoma (TCGA-LIHC) cohort. Additionally, we identified CCCTC-binding factor (CTCF) as a transcription activator of TFAP4 gene transcription. Our findings suggest that TFAP4 directly regulates TERT gene transcription via CTCF in non-HBV associated HCCs.
To dissect the transcriptional regulation of TERT in human HCC, we performed dual luciferase reporter assay. We used the wild-type TERT promoter [10] and an overexpression approach, cloning a series of transcription factors suggested to alter TERT promoter activity. We then subjected these factors to reporter assay using Huh7 and PLC/PRF/5 HCC cells. We screened a total of 25 transcription factors and found that eight of them significantly induced the wild-type TERT promoter activity upon overexpression of the transcription factors compared to the empty vector control in both Huh7 and PLC/PRF/5 cells (Figure 1A and 1B). To further analyze the correlation between the transcription factors and TERT promoter activity in the context of HCC, we performed correlation analysis of their mRNA expression using TCGA-LIHC cohort. From this analysis, we found that TFAP4 and MYBL2 were significantly associated with the TERT mRNA expression in TCGA-LIHC cohort (Figure 1C).

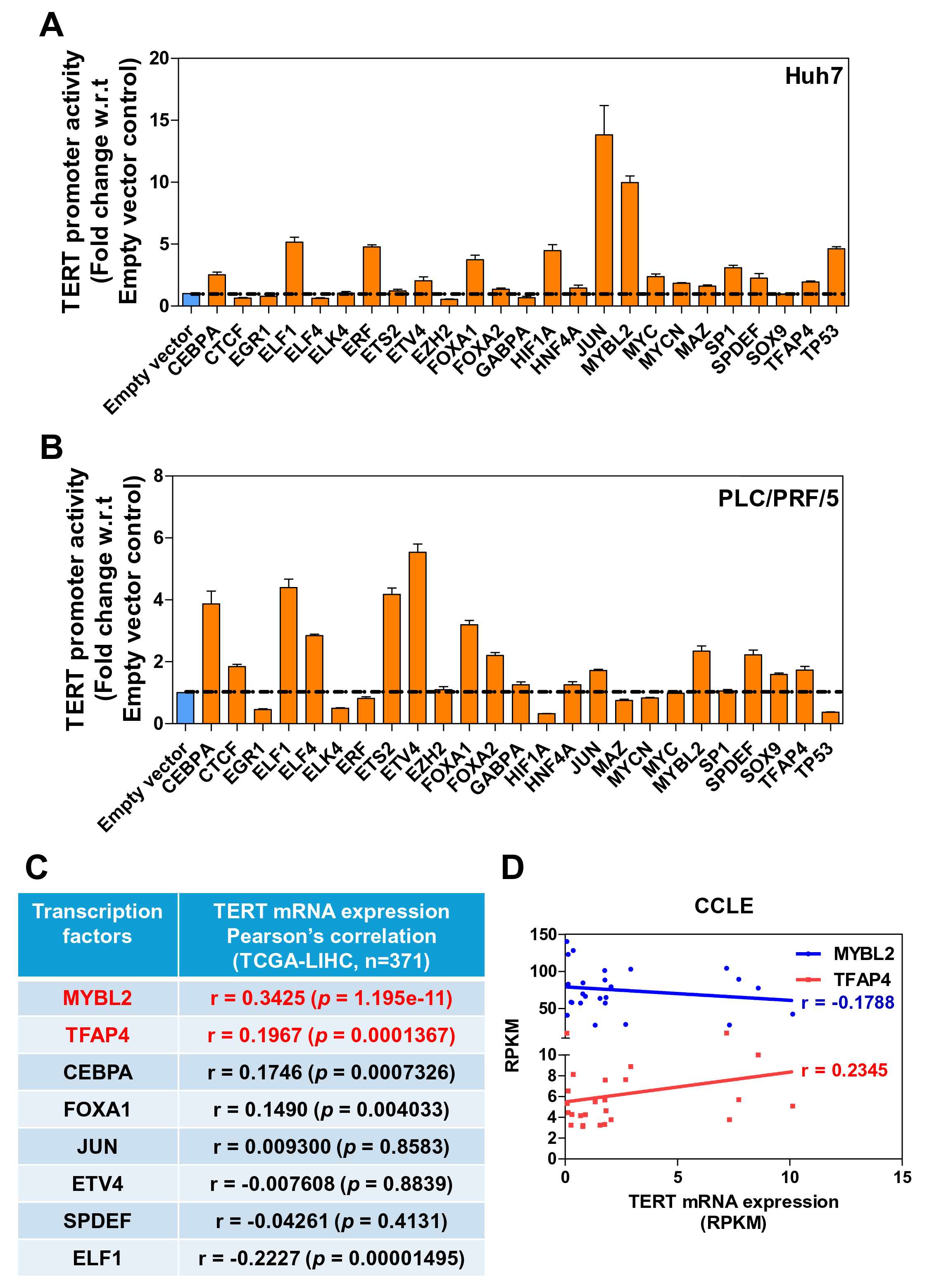
Figure 1. Screening of transcription factors governing TERT mRNA expression in human HCC. (A-B) Overexpression approach was used for screening transcription factors that might govern TERT mRNA transcription in Huh 7 (A) and PLC/PRF/5 cells (B) by dual luciferase reporter assay. (C) Correlation analysis between potential transcription factors and TERT mRNA expression in human HCC in TCGA-LIHC cohort (n=371). (D) Correlation analysis between MYBL2, TFAP4, and TERT mRNA expression in the Cancer Cell Line Encyclopedia (CCLE) liver dataset (ns=P>0.05, *P<0.05, **P<0.001, ***P<0.001).
To further assess the correlation between transcription factor and TERT mRNA expression in the context of liver cancer cell lines, we conducted a correlation analysis between TFAP4, MYBL2, and TERT mRNA expression using the Cancer Cell Line Encyclopedia (CCLE). We observed that TFAP4 was significantly and positively associated with the TERT mRNA expression in the liver cancer cell lines in the CCLE dataset, while MYBL2 was negatively associated with the TERT mRNA expression. The findings suggest that TFAP4 is a potential candidate governing TERT mRNA transcription in HCC (Figure 1D).
We used TIMER (Tumor Immune Estimation Resource) [15] to determine the TFAP4 expression levels in different tumor types from TCGA database. Differential gene expression analysis showed that TFAP4 was upregulated in tumors compared to their corresponding non-tumor tissues in most cancer types, including liver cancer (P<0.001) (Figure 2A). We used TCGA RNA-sequencing database to analyze the expression of the TFAP4 gene in HCC tumors. The boxplots showing the association of gene expression and tumor stage revealed that the TFAP4 mRNA expression levels in HCC tumors (n=371), which include all etiologies, were upregulated compared with the corresponding non-tumor liver samples (Figure 2B). Similarly, TERT mRNA expression levels in tumor tissues (n=371) were upregulated compared with the corresponding non-tumor liver samples (Figure 2C). Additionally, the correlation analysis suggests that the mRNA expression of TFAP4 is significantly associated with the TERT mRNA expression in TCGA-LIHC cohort (Figure 2D). Intriguingly, we found that TFAP4 mRNA expression was not significantly associated with TERT mRNA expression in our HBV-associated HCC cohort consisting of 41 pairs of HCC (P=0.2168) (Figure 2E). Furthermore, in TCGA-LIHC cohort, patients with high mRNA expression of either TFAP4 or both TFAP4 and TERT had significantly reduced overall survival rates. However, such correlation was not observed in our HBV-associated HCC cohort (Figure S1). It is of note that TCGA cohort included HCC of all etiologies, i.e., both non-HBV infection and HBV infection, while our HCC cohort included only HBV-associated HCC. Hence, these discrepancies suggest that the positive association between TFAP4 and TERT mRNA expression may be independent of HBV infection or HBV viral integrations.
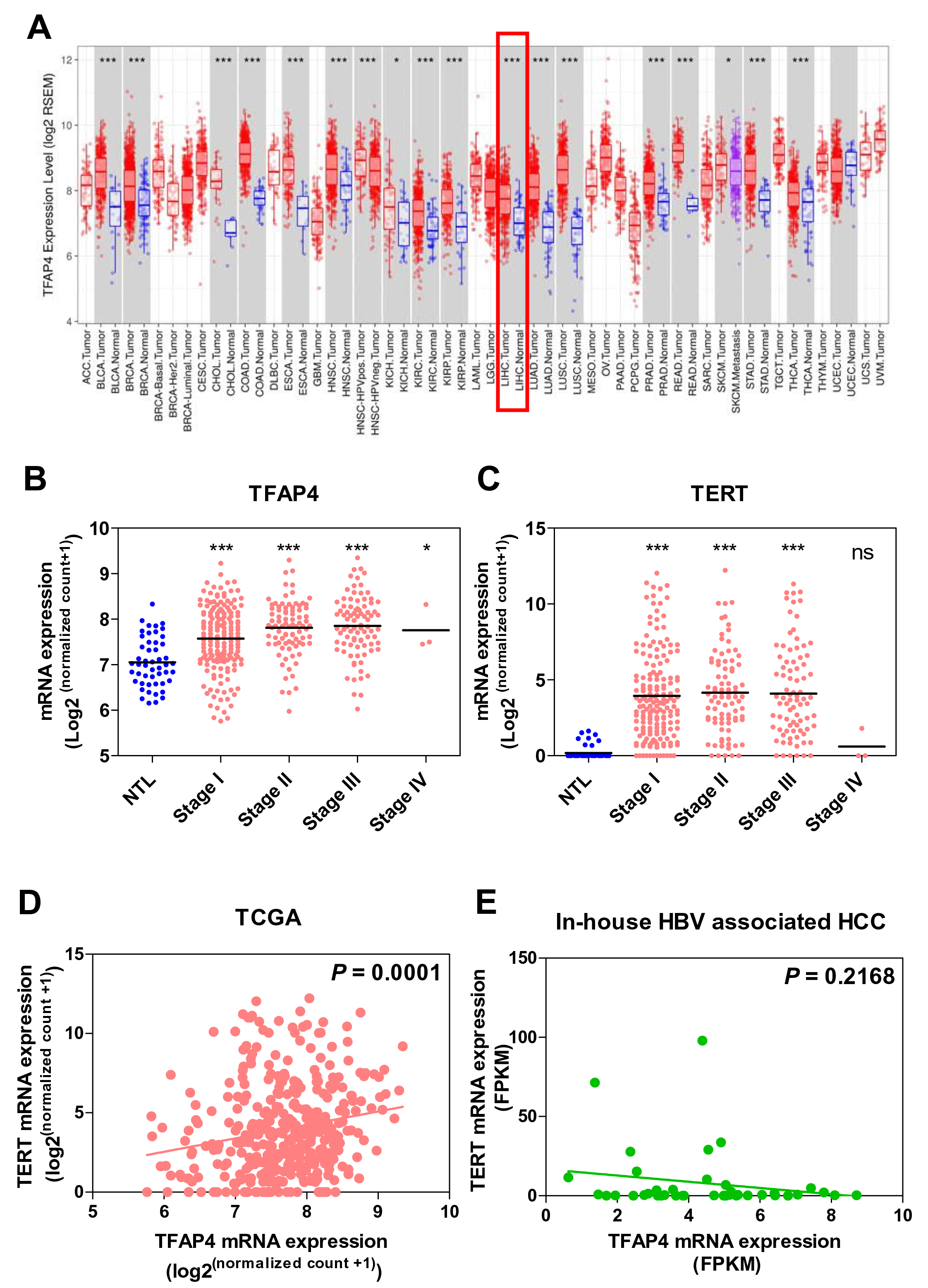
Figure 2. TFAP4 is up-regulated in human HCC and significantly correlates with TERT mRNA expression in TCGA-LIHC cohort. (A) TFAP4 expression levels in different tumor types from TCGA database were determined by TIMER. The distributions of gene expression levels are displayed, with statistical significance indicated. TFAP4 is upregulated in tumors compared to their corresponding non-tumor tissues in almost all cancer types, including liver cancer. The red rectangle indicates the expression levels in liver cancer. (B) A tumor stage plot of TFAP4 using TCGA-LIHC database showing an upregulation of TFAP4 mRNA levels in tumors compared to the corresponding non-tumor livers. (RPKM: reads per kilobase million) (C) A tumor stage plot of TERT using TCGA-LIHC database showing an upregulation of TERT mRNA levels in tumors compared to the corresponding non-tumor liver samples. (D) Correlation between TFAP4 and TERT in human HCCs in TCGA-LIHC cohort. (E) Correlation between TFAP4 and TERT in human HCCs in in-house HBV-associated HCC cohort (ns = P>0.05, *P<0.05, **P<0.001, ***P<0.001).
To further investigate the role of TFAP4 in TERT gene transcription in HCC, we examined both TFAP4 protein expression and telomere length in HCC cell lines. We found that TFAP4 protein was ubiquitously expressed in most of the HCC cell lines tested as well as the immortalized normal liver cell line MIHA (Figure 3A). With qRT-PCR used to measure the telomere length, Huh7 and Hep3B cells had relatively more telomere copies compared to other HCC cell lines and MIHA cells (Figure 3B). Therefore, we selected Huh7 and Hep3B cells for subsequent functional validations. To study the functional role of TFAP4 in human HCC in vitro, we established TFAP4 stable knockdown in these two cell lines, using shRNA targeting lentiviral expression system. We examined the TERT mRNA expression and relative telomere copy numbers. Upon TFAP4 knockdown in Huh7 cells, there was a significant reduction in both TERT mRNA expression and relative telomere copy numbers compared to non-target control (NTC) cells (Figure 3C). In contrast, upon TFAP4 knockdown in Hep3B cells, which are HBV-infected HCC cells [16], both the TERT mRNA expression and relative telomere copy numbers were not significantly altered (Figure 3D).
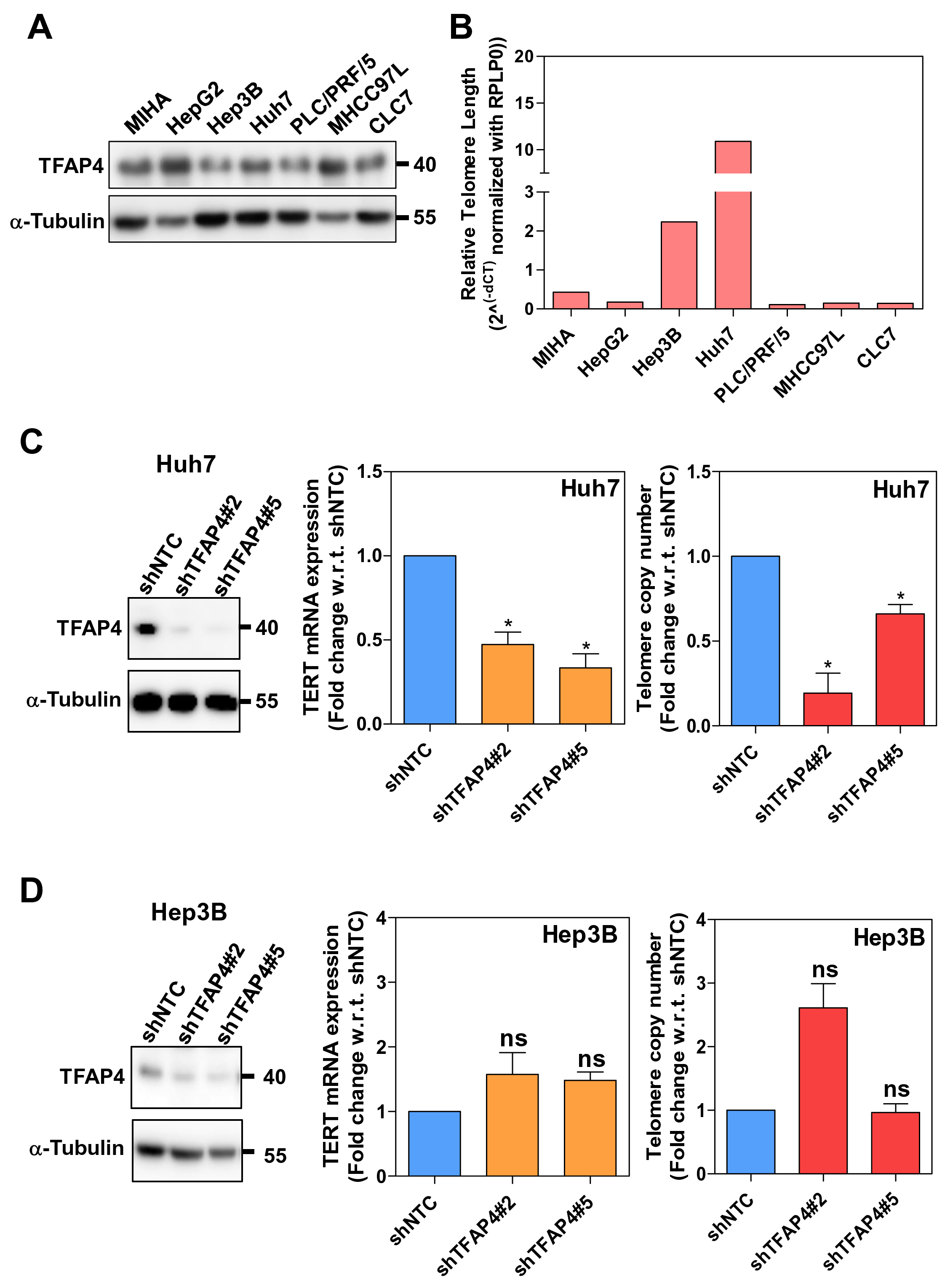
Figure 3. TFAP4 is a direct transcription factor driving telomerase reactivation in non-HBV-associated HCCs. (A) The protein expression levels of TFAP4 in HCC cell lines. α-Tubulin served as internal control in western blot analysis. (B) The relative telomere length in HCC cell lines measured by qRT-PCR. RPLP0 was used as a normalizer for DNA input in this measurement. (C) Establishment of TFAP4 stable knockdown clones in Huh7 cells. TERT mRNA expression and relative telomere copy numbers were significantly reduced upon TFAP4 knockdown. (D) Establishment of TFAP4 stable knockdown clones in Hep3B cells. TERT mRNA expression and relative telomere copy numbers were not significantly impacted upon TFAP4 knockdown (ns = P>0.05, *P<0.05).
Regarding the cellular functional significance of TFAP4 in HCC cells, we observed that in shTFAP4 Huh7 cells, the cell invasive ability was significantly reduced as compared with NTC (Figure S2A). However, the shTFAP4 Huh7 cells showed a significant reduction of migratory ability only with sequence shTFAP4#5 (Figure S2A). In shTFAP4 Hep3B cells, only sequence shTFAP4#5 but not shTFAP4#2 showed a significant reduction in migratory and invasive abilities, as compared with NTC (Figure S2B). Taking together, the findings suggest that presence of both TFAP4 and TERT mRNA expression may contribute to cell invasiveness in Huh7 cells (in which TERT mRNA was altered), while single knockdown of TFAP4 without TERT reduction as in Hep3B cells (in which TERT mRNA was not significantly altered), such cellular function was diminished.
Using the INSECT (In-silico Search Co-occurring Transcription Factors) 2.0 webserver, we identified the putative binding sites of TFAP4 on the TERT promoter region. We analyzed the score threshold of TFAP4 occurrence on the TERT promoter and chose the four positions (–327, –286, –234, and –111nt) with relatively higher scores (Figure 4A). To assess the binding of TFAP4 on the TERT promoter region, we performed dual luciferase reporter assay. The results showed that upon mutation at positions –327, –234, and –111nt, there was a significant decrease in promoter activity compared to Huh7 cells transfected with the wild-type TERT promoter (Figure 4B). Similarly, in HepG2 cells transfected with the wild-type TERT promoter, any mutation at the putative TFAP4 binding site among the four suggested positions significantly decreased promoter activity (Figure 4C). Notably, when the putative TFAP4 binding site at the –111nt position was mutated, the promoter activities drastically decreased in both Huh7 and HepG2 cells (reduced by 69.4% and 90%, respectively) (Figure 4B and 4C). Finally, with ChIP-qPCR assay using the TFAP4 antibody, we demonstrated that TFAP4 physically interacted with the TERT promoter at the –111nt position, but not at either the –234 or –286nt position in Huh7 cells (Figure 4D). In summary, TFAP4 is a direct transcription factor which physically interacts with the TERT promoter region and drives telomerase reactivation in non-HBV-associated HCC cells.
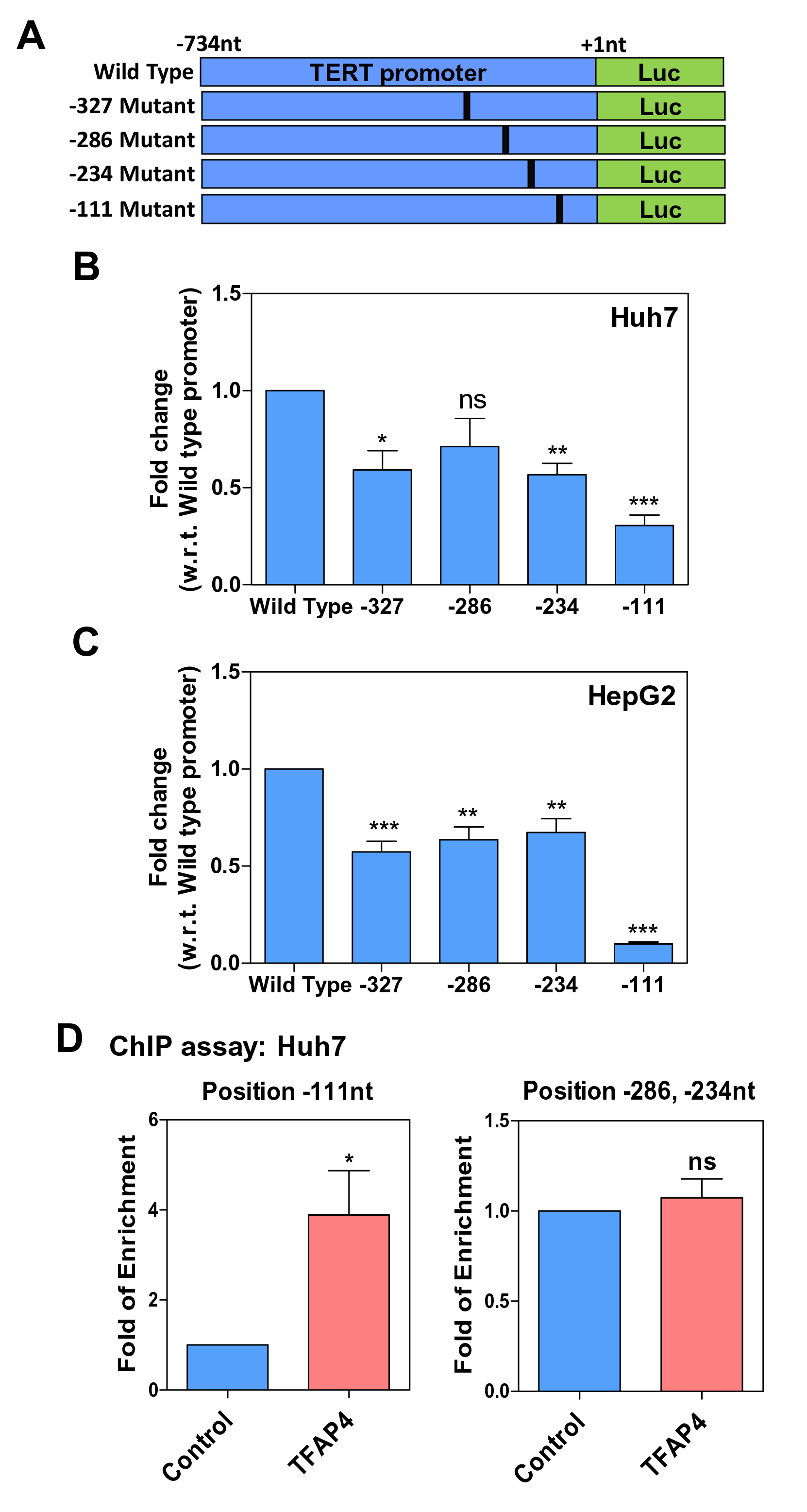
Figure 4. TFAP4 as a transcriptional regulator of TERT in HCC cell line. (A) A schematic diagram of the wild-type TERT promoter and mutations of the four putative transcription factor binding motifs at –327, –286, –234, and –111nt in the TERT promoter. (B) Dual luciferase reporter assays of TERT promoters of wild type and with the corresponding mutations in Huh7 cells are shown. (C) Results of dual luciferase reporter assays of TERT promoters of wild type and with the corresponding mutations in HepG2 cells. (D) ChIP assays using specific antibodies against TFAP4 in Huh7 cells. The results confirmed that TFAP4 specifically bound to and were enriched on –111nt but not –286 and –234nt of the TERT promoter region (ns = P>0.05, *P<0.05, **P<0.01, ***P<0.001).
While TFAP4 is upregulated in HCC, its gene transcription regulation control in HCC remains unknown. To investigate the transcription regulator of TFAP4, we searched for putative DNA binding proteins from the Encyclopedia of DNA Elements (ENCODE) data portal [17] in the context of HepG2 cells and liver tissue. According to the ChIP-sequencing database from ENCODE data portal, CTCF, FOXA2, MYC, and MAZ are suggested to physically interact with the TFAP4 promoter region. Using dual luciferase reporter assay, we examined the transcription activity among the four putative transcription factors on the TFAP4 promoter by using an overexpression approach in Huh7 cells. Only CTCF, FOXA2, and MAZ significantly increased TFAP4 promoter activity (Figure 5A). To further explore which transcription factors drove TFAP4 gene transcription in HCC, we conducted a correlation analysis among CTCF, MAZ, and TFAP4 mRNA expression using the Cancer Cell Line Encyclopedia (CCLE). The correlation analysis suggested that both CTCF and MAZ were significantly associated with TFAP4 mRNA expression in liver cancer cell lines in the CCLE dataset, with CTCF having the most significant positive correlation compared to MAZ. This suggests that CTCF is one of the potential candidates governing TFAP4 mRNA transcription in HCC (Figure 5B). Moreover, upon CTCF knockdown in Huh7 cells, both TFAP4 and TERT showed a significant reduction at the mRNA and protein expression level compared with shNTC (Figure 5C). In addition, CTCF knockdown led to significant reductions in relative telomere copy number compared with shNTC Huh7 cells (Figure 5D). Finally, with immunohistochemistry, we found that CTCF protein is overexpressed in human HCC compared to corresponding non-tumor counterparts (Figure 5E), suggesting that CTCF protein is overexpressed in human HCC and facilitates TFAP4 gene transcription.
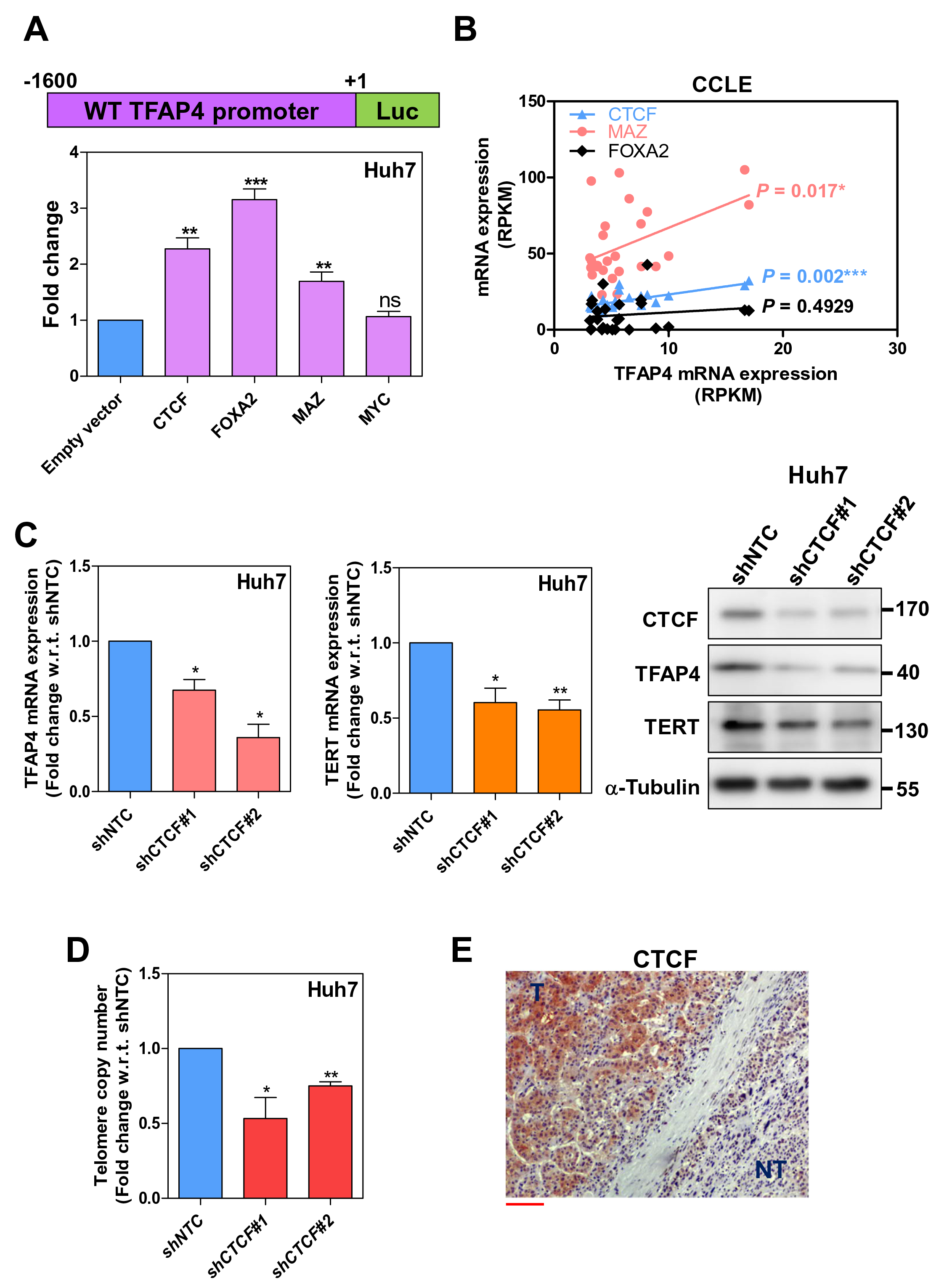
Figure 5. CTCF is crucial for TFAP4 gene transcription in human HCCs. (A) A schematic diagram of the wild-type TFAP4 promoter (upper panel). Results of dual luciferase reporter assays of wild type TFAP4 promoter with overexpression of CTCF, FOXA2, MAZ, and MYC in Huh7 cells are shown (lower panel). (B) Correlation analysis between CTCF, MAZ, and TFAP4 mRNA expression in the CCLE dataset. (C) TFAP4 and TERT mRNA (left panel) and protein expression (right panel) upon CTCF knockdown in Huh7 cells. (D) The relative telomere copy numbers upon knockdown of CTCF compared with non-target control in Huh7 cells. (E) A representative example of immunohistochemical staining for CTCF protein in human HCCs. ‘T’, tumor; ‘NT’, non-tumor liver. Scale bar, 100 μm (ns = P>0.05, *P<0.05, **P<0.01).
To further dissect the possible mechanism underlying the positive correlation between TFAP4 and TERT mRNA expression in TCGA cohort which includes HCCs of all etiologies, but not in our in-house HBV-associated HCCs, we further stratified the patients in TCGA cohort according to the etiologies for analysis. Interestingly, a significant correlation between TFAP4 and TERT mRNA expression was observed only in patients with chronic alcohol consumption, but not those with either HBV- or HCV-associated HCC (Figure 6A and Figure S3).

Figure 6. Ethanol enhances both TFAP4 and TERT mRNA expression in human non-HBV-associated HCCs and non-HBV-infected HCC cells. (A) Correlation analysis between TRAP4 and TERT mRNA expression levels in human HCCs related to chronic alcoholism (n=72, left panel) and HBV-associated HCC (n=76, right panel) in TCGA-LIHC cohort. (B) TFAP4 and TERT mRNA expression upon ethanol treatment at 50 mM for 24 hours in HepG2 cells (left 2 panels) and Hep3B (right 2 panels). (C) Methylation status at –96nt of the TERT promoter region upon ethanol treatment in HepG2 and Hep3B cells. (D) Dual luciferase reporter assays of wild-type 6x TFAP4 consensus DNA binding motif-containing promoter, with or without 50 mM ethanol treatment in HepG2 and Hep3B cells. Empty vector (EV) with no treatment served as normalization control (ns = P>0.05, *P<0.05, **P<0.01).
Therefore, we conducted in vitro experiments on HCC cells with or without HBV infection. We treated HepG2 and Hep3B cells with 50 mM ethanol for 24 hours. We observed a significant up-regulation of both TFAP4 and TERT mRNA expression in HepG2 cells upon ethanol treatment (Figure 6B), while there was no significant impact on TFAP4 mRNA expression in Hep3B cells. In contrast, the TERT mRNA expression was significantly reduced in Hep3B cells (Figure 6B). Given that it has been reported that ethanol might alter the methylation status using a rat model [18], we further investigated the methylation status of the TERT promoter region upon ethanol treatment. From above finding (Figure 4) suggested that proximity of transcription start site of TERT promoter region crucial for TFAP4 binding (–111nt). To further investigate the methylation status of this region, we performed in-silico analysis of the CpG methylation site within this region and found that -96nt is the putative CpG methylation site, which is in close proximity of the TFAP4 binding site in the TERT promoter. Using methylation-specific qPCR targeting the TERT promoter region, we found there was an induction of DNA methylation status at the TERT promoter region upon ethanol treatment in Hep3B, but not HepG2 cells (Figure 6C). To further assess whether ethanol would alter TFAP4 transcription activity in HCC cells, we employed the six times repeat TFAP4 consensus DNA binding sequence (encode nucleotide sequence “CAGCTG”) driven luciferase reporter constructs to conduct Dual luciferase reporter assay to assess TFAP4 transcription activity upon ethanol treatment. With ethanol treatment, there were no significant alterations of TFAP4 transcription activity in both HepG2 and Hep3B cells (Figure 6D). The findings suggest that ethanol may induce TFAP4 and TERT mRNA expression in HCC cells and alcohol-related HCC.
Telomerase reactivation has been extensively investigated over decades in cancer development and stem cell regeneration [19, 20]. TERT promoter mutations are the most frequently reported genetic alteration at the TERT locus in various types of cancer [19]. Recent studies discovered that other TERT promoter mutations such as -G124A and -G146A create new ETS DNA binding sites, enhancing TERT reactivation in various cancer types including HCC [21, 22]. This inspires scientists to use the TERT promoter as a serum cell-free DNA diagnostic marker for metabolic-dysfunction-associated steatotic liver disease (MASLD)-associated HCC [23, 24]. In HCC, apart from promoter mutations, HBV DNA integration at the TERT promoter locus also plays a crucial role in driving TERT mRNA expression, serving as an actionable therapeutic target for antisense oligonucleotides-mediated HCC treatment [5]. Even though genetic alteration contributes to most of the TERT gene transcription in HCC, a significant portion of HCC with TERT reactivation occurs without genetic alteration [10], leading us to investigate the underlying mechanism of TERT reactivation without genetic alteration in human HCC.
In this study, we identified TFAP4 as a direct TERT reactivation factor and is under the transcriptional control of CTCF in human HCC. TFAP4 was first identified as a c-MYC downstream regulator in colorectal and breast cancer and plays a role in cell cycle control and metastasis in cancer development [25–27]. Later, MYCN was suggested to be one of the potential transcriptional regulators of TFAP4 in neuroblastoma [28, 29]. In HCC, TFAP4 has been documented to activate the Wnt//-catenin and PI3K/AKT signaling pathways, which play vital roles in tumor initiation and promoting metastasis [30, 31]. In this study, we further expanded on this understanding by revealing that TFAP4 is instrumental in the reactivation of TERT in HCC cells. This reactivation would in turn aid in tumor initiation and progression by maintaining the stability of the telomere. In this study, TFAP4 was also expressed in MIHA cells, which is an immortalized normal liver cell line (Figure 3A). According to a recent study on pluripotency factor-induced reprogramming of mouse fibroblasts and hepatocytes [32], during the iPSC reprogramming, Tfap4 was significantly up-regulated. Conversely, upon co-knockdown of Tfap4 with Irf6, the reprogramming efficiency was diminished. These findings may suggest that, during the establishment of immortalized normal liver cell line, TFAP4 is required to participate in this process. However, whether protein expression level of TFAP4 has any effects on MIHA cells remains to be elucidated.
CTCF protein was first identified in chicken in 1990 [33] and is a highly conserved zinc finger protein. It serves as a chromatin modifier and X-inactivation factor in cells [34, 35]. CTCF has been suggested to negatively regulate TERT mRNA expression via its binding to the proximal exonic region of TERT [36]. In HCC, the loss of chromosome 16q22.1, where CTCF gene is located, is implicated with an increase in intrahepatic HCC formation [37], while a contradictory suggestion proposes that CTCF cooperates with FOXM1 to facilitate tumor growth and promote metastasis in HCC [38]. Moreover, CTCF also plays a role in enhancing stem cell-like properties in gastric cancer [39]. In this study, we showed that CTCF was overexpressed and directly activated TFAP4 gene transcription, resulting in the activation of TERT mRNA expression in HCC cells. Previous studies suggest that CTCF directly acts on TERT mRNA expression in lung cancer [40], suggesting that the transcription activity and functional role of CTCF on the TERT locus may be context-dependent. Further studies on the role of CTCF in HCC are warranted.
Concerning DNA methylation, DNA hypomethylation promoting TERT mRNA expression has been documented in cervical cancer [41], while hypermethylation suppresses TERT mRNA expression in esophageal squamous cell carcinoma [42]. Recently, Lee and colleagues summarize the TERT promoter methylation status in human cancers, revealing that both hypomethylation and hypermethylation (dual-methylation) can be present within different regions of the TERT promoter in cancer cell lines. This suggests that hypomethylation close to the transcription start site might serve as a switch for TERT promoter activation [43]. Apart from intrinsic methylation in cells, external factors governing DNA methylation also play an important role in gene expression, such as ethanol [44–46]. Interestingly, in HCC patients with chronic alcohol consumption, there is a significant correlation between TERT and TFAP4 mRNA expression in their tumors, and this finding aligns with our experimental validation in vitro and in TCGA-LIHC cohort. This further supports the idea that ethanol might be one of the factors inducing HCC formation. However, we could not detect changes in the methylation status at the TERT promoter locus in HepG2 cells upon ethanol treatment. Whether there are other alterations on TFPA4 influencing TERT reactivation in non-viral associated HCC remains to be elucidated in future studies. Lastly, given the recent global rise in the incidence of MASLD-associated HCC, it will be interesting to examine whether CTCF or TFAP4 plays a role in these HCCs.
In summary, our findings demonstrated that TFAP4 directly regulates TERT gene transcription via CTCF in non-HBV-associated HCCs, but its role in regulating telomerase expression or activity through HBV DNA integration in HBV-associated cells may be limited.
We randomly selected HCC cases from patients who underwent surgical resection of the tumors at Queen Mary Hospital, Hong Kong. Following surgical resection, all specimens were immediately snap frozen in liquid nitrogen and stored at -80°C, with informed consents from the patients. The use of human clinical samples was approved by the Institutional Review Board of the University of Hong Kong and the Hong Kong Hospital Authority Hong Kong West Cluster (UW 17-056).
Human HCC cells, HepG2, Hep3B and PLC/PRF/5 were obtained from American Type Culture Collection. Huh7 were purchased from the Japanese Collection of Research Bioresources Cell Bank. Human HCC cell line MHCC97L was a gift from Dr. ZY Tang (Fudan University, Shanghai, China) and the STR authentication was conducted. Immortalized liver cell line MIHA was obtained from Dr. J Roy-Chowdhury (Albert Einstein College of Medicine, New York, USA). CLC7 was a gift from Dr. Lijian Hui (Shanghai Institutes for Biological Sciences, CAS, Shanghai, China). Human embryonic kidney cell line 293FT was obtained from Invitrogen (Carlsbad, USA). HepG2, Hep3B and PLC/PRF/5 were cultured in Minimum Essential Medium containing 10% fetal bovine serum and 1mM sodium pyruvate, CLC7 was cultured in RPMI-1640 Medium containing 10% fetal bovine serum, while the others were cultured in Dulbecco’s modified Eagle’s medium high glucose supplemented with 10% fetal bovine serum.
We amplified full-length forms of transcription factors from HepG2 complementary DNA (cDNA) and subcloned them into the pcDNA3.1+neo vector. We amplified the TERT promoter (chr 5. 52,411,030-52,410,031) and TFAP4 promoter (chr 16. 4,274,618-4,256,186) from HepG2 genomic DNA and subcloned them into the pGL3-Basic vector. We also amplified a series of TERT or TFAP4 promoter DNA fragments with mutations from the wild-type TERT or TFAP4 promoter/pGL3-Basic vector and sub-cloned them into the empty vector of pGL3-Basic. We purchased short-hairpin (sh) RNA interfering sequences targeting CTCF and TFAP4 from Horizon Discovery (Cambridge, United Kingdom) and synthesized them by Integrated DNA Technologies (Coralville, IA, USA), before subcloning them into the pLKO.1-Puro vector. The primers used for cloning are listed in Table S1.
Huh7 and PLC/PRF/5 cells underwent transfection with various combinations of plasmids using Lipofectamine 3000 (Invitrogen) as per the manufacturer’s protocol. The plasmids included TERT wild-type promoter/pGL3-Basic, TFAP4 wild-type promoter/pGL3-Basic, different forms of mutated TERT and TFAP4 promoter/pGL3-Basic reporter constructs, and an internal control pRL/PGK. After 24 hours of transfection, we measured luciferase and Renilla luciferase activities using a Dual Luciferase Reporter assay system (Promega, Madison, WI, USA) according to the manufacturer’s protocol. The Renilla luciferase activity was used to normalize transfection efficiency. We conducted these experiments independently at least three times.
We employed a lentiviral-mediated approach to construct stable CTCF and TFAP4 knockdown HCC cell lines. We purchased human ON-TARGETplus SMARTpool siRNA duplexes, which target CTCF and TFAP4, along with a non-target control from Horizon Discovery. We stably transduced the pLKO.1-Puro vectors, which contain shRNAs targeting corresponding gene targets and a non-target control, into HCC cell lines. We performed puromycin selection to achieve the stable expression of shRNAs and shNTCs. The sequences of all shRNAs are listed in Table S1.
Cells were lyzed in NET (150 mM NaCl, 5 mM EDTA, 50 mM Tris-HCl, pH8.0) buffer with 1%NP-40 and equal amounts of protein were separated in sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis gel for western blot analysis. Immunodetection was performed using antibodies which are listed in Table S2.
Total RNA was extracted from HCC cell lines using Trizol reagent (Invitrogen). The cDNA synthesis using GeneAmp PCR Reagent Kit (Applied Biosystems, Foster City, CA, USA) was used to reverse messenger RNA into cDNA and ABI Power SYBR™ Green PCR Master Mix (Applied Biosystems) coupled with primer specific for TERT, TFAP4 and HPRT1 were used for measurement. The primer sequences are listed in Table S3.
The detailed protocol has been described [47]. The antibody against TFAP4 protein (HPA001912, Sigma-Aldrich) and normal Mouse IgG antibody (sc-3877, Santa Cruz biotechnology) were used for the chromatin immunoprecipitation (ChIP) assay. The primers (–127nt to +1nt: 5’-CCGCCTCCTCCGCGCGGACC-3’ and 5’-CAGCGCTGCCTGAAACTC-3’; –286nt to –234nt: 5’-GGGAGCGCG AGCGGCGCGCGGG-3’ and 5’-CGACAGCGCAGCTGCTCCGGGC-3’) covering the putative TFAP4 binding site on TERT DNA region were employed for standard qRT-PCR using ABI Sybr green Gene Expression Assay supplements of AmpliTaq Gold 360 master mix (Applied Biosystems) and detected by ABI QuantStudio 5 Real-Time PCR System (Applied Biosystems) in the ChIP assay.
We seeded 1,000 cells on each well of a 96-well plate and cultured them for 5 days. Every 24 hours (Day 1 to Day 5), we fixed designated cells using 30 μL of 100% methanol per well for 5 minutes, and then replaced it with 100 μL/well of PBS after cell fixation. After 5 days, we removed the PBS and used diluted DAPI at 1 μg/mL to stain the cell nuclei. We then subjected the plate to fluorescent photo capturing and cell counting by the ImageXpress Pico Automated cell Imaging System (Molecular Devices, San Jose, CA, USA). The cell counting program setting was pre-configured by the manufacturer. We conducted these experiments independently at least three times.
100,000 cells were resuspended into 100 μL of serum-free medium and added to the upper chamber of the insert with 8 μm pore size membrane (Merck Millipore, Burlington, MA, USA). For the invasion assay, the transwell was pre-coated with cold 1:4 diluted matrix gel and incubated at 37 C for 1 hour before use. For the bottom chamber, 500 CL of standard culture medium was added to the bottom chamber of cell culture insert in the well of 24-well plate acting as chemoattractant. After incubation, cells on the insert membrane were fixed by 100% methanol for 5 minutes and stained with crystal violet solution for 20 minutes. After washing the insert with water, non-migrated cells on the upper side of the membrane were gently removed by cotton swab. Photos were taken from three randomly selected fields with migrated cells on the lower side of the membrane and cells counted manually. Experiments were done at least three independently.
We quantified the relative telomere length using quantitative real-time PCR (qRT-PCR), according to the method described by O’Callaghan and Fenech in 2011 [48] with minor modifications. Firstly, cells were centrifuged for 5 minutes at 300 x g. After discarding the supernatant, we re-suspended the cell pellet in 200 μL PBS. Next, we isolated the DNA using the TIANamp Genomic DNA Kit (Tiangen Biotech Co., Ltd., Beijing, China), as per the manufacturer’s instructions. We used these samples for standard qRT-PCR using the ABI Sybr Gene Expression Assay (Applied Biosystems) and detected them using the ABI QuantStudio 5 Real-Time PCR System (Applied Biosystems). The 36B4 gene, which encodes an acidic ribosomal protein lateral stalk subunit P0 (RPLP0), served as internal control. The qRT-PCR primers used are listed in Table S4. We performed all reactions in triplicate.
We isolated genomic DNA from HCC cell lines using the TIANamp Genomic DNA Kit (Tiangen Biotech Co., Ltd., Beijing, China) according to the manufacturer’s instructions. To perform methylation-specific quantitative PCR, we subjected 1 μg of genomic DNA to methylation-sensitive restrictions enzyme Hha I digestion at 37 °C for three hours. We then subjected 50 ηg of digested genomic DNA to standard quantitative PCR using the ABI Sybr Gene Expression Assay (Applied Biosystems) and detected them using the ABI QuantStudio 5 Real-Time PCR System (Applied Biosystems). The primers (–127nt to +1nt: 5’-CCGCCTCCTCCGCGC GGACC-3’ and 5’-CAGCGCTGCCTGAAACTC-3’) covering the putative methylation site on TERT DNA region were employed for standard qPCR in this assay. The same amount of undigested genomic DNA served as input DNA control.
We analyzed the data using the Prism 5 software (GraphPad Software Inc., San Diego, CA, USA). Depending on the nature of the study, we subjected the data to either paired or unpaired t-test. We used paired t-test to compare gene expressions in HCC tumor tissues with corresponding non-tumor liver samples, as well as to analyze gene expression changes in independent samples via qRT-PCR. For Dual luciferase reporter assay analyses, we employed paired t-test. In contrast, we used non-linear regression test (curve fit) to analyze cell proliferation assay data. We considered test results significant when the P-value was less than 0.05. For vitro functional assays, we expressed data as mean ± SEM. We defined statistical significance as *P<0.05, **P<0.01, and ***P<0.001.
The following supplementary materials are available: CHP2401020008SupplementaryMaterials.zip.
Figure S1. Survival analysis upon overexpression of TFAP4 mRNA in human HCC.
Figure S2. HCC cell proliferation, migratory ability and cell invasiveness upon knockdown of TFAP4.
Figure S3. TFAP4 mRNA expression is not significantly correlated with TERT mRNA expression in HCV-associated HCC.
Table S1. Primers used for cloning plasmids.
Table S2. Antibodies used in this study.
Table S3. Primers used for qRT-PCR.
Table S4 Primers used for telomere length measurement.
The use of human clinical samples was approved by the Institutional Review Board of the University of Hong Kong and the Hong Kong Hospital Authority Hong Kong West Cluster (UW 17-056).
Not applicable.
The original data presented in the study are openly available in UCSC Xena browser at (https://xenabrowser.net/).
The study was supported by the Hong Kong Research Grants Council (RGC) Theme-based Research Scheme (T12-704/16-R and T12-716/22-R), Innovation and Technology Commission grant to the State Key Laboratory of Liver Research of the University of Hong Kong (ITC PD/17-9), University Development Fund of The University of Hong Kong. I.O.L. Ng is Loke Yew Professor in Pathology.
The authors have declared that no conflict of interest exists.
Conceptualization: K.M.F.S. and I.O.N.; Methodology: H.L., T.K.H.W., K.M.F.S. and I.O.N.; Investigation: H.L., T.K.H.W., K.M.F.S., E.M.S.S., T.C.Y.Y., J.M.F.L., Y.M.T. and D.W.H.H.; Resources: I.O.N; Writing: H.L., T.K.H.W., K.M.F.S. and I.O.N.; Supervision: K.M.F.S. and I.O.N.; Visualization: K.M.F.S. and I.O.N.; Project administration: K.M.F.S. and I.O.N.; Funding acquisition: I.O.N.
complementary DNA Chromatin immunoprecipitation CCCTC-binding factor; 4',6-diamidino-2-phenylindole; E74 like ETS transcription factor 4; E26 transformation-specific; fluorescein isothiocyanate; Fragments Per Kilobase Million; Hepatocellular carcinoma; Hepatitis B Virus; Hepatitis C Virus; hypoxia inducible factors; Jun proto-oncogene, AP-1 transcription factor subunit; MYC proto-oncogene, bHLH transcription factor; nuclear factor kappa B subunit 1; Paired Box Proteins; quantitative Real Time Polymerase Chain Reaction; sodium dodecyl sulfate; short tandem repeat; Sp1 transcription factor; signal transducer and activator of transcription 3; signal transducer and activator of transcription 5; The Cancer Genome Atlas Liver Hepatocellular Carcinoma cohort; Telomerase Reverse Transcriptase; Transcription Factor AP-4.
| 1. | Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021;71(3):209-249. [Google Scholar] [CrossRef] |
| 2. | Ozakyol A. Global Epidemiology of Hepatocellular Carcinoma (HCC Epidemiology). J Gastrointest Cancer. 2017;48(3):238-240. [Google Scholar] [CrossRef] |
| 3. | Jiang Y, Chen C, Chen SM, Wang YQ, Xu Y, Wang Y, et al. Telomerase reverse transcriptase promotes the proliferation of human laryngeal carcinoma cells through activation of the activator protein 1. Oncol Lett. 2013;6(1):75-80. [Google Scholar] [CrossRef] |
| 4. | Autexier C, Lue NF. The structure and function of telomerase reverse transcriptase. Annu Rev Biochem. 2006;75:493-517. [Google Scholar] [CrossRef] |
| 5. | Ningarhari M, Caruso S, Hirsch TZ, Bayard Q, Franconi A, Vedie AL, et al. Telomere length is key to hepatocellular carcinoma diversity and telomerase addiction is an actionable therapeutic target. J Hepatol. 2021;74(5):1155-1166. [Google Scholar] [CrossRef] |
| 6. | Dogan F, Forsyth NR. Telomerase Regulation: A Role for Epigenetics. Cancers (Basel). 2021;13(6):1213. [Google Scholar] [CrossRef] |
| 7. | Zhao Y, Cheng D, Wang S, Zhu J. Dual roles of c-Myc in the regulation of hTERT gene. Nucleic Acids Res. 2014;42(16):10385-10398. [Google Scholar] [CrossRef] |
| 8. | Khattar E, Tergaonkar V. Transcriptional Regulation of Telomerase Reverse Transcriptase (TERT) by MYC. Front Cell Dev Biol. 2017;5:1. [Google Scholar] [CrossRef] |
| 9. | Cong YS, Wen J, Bacchetti S. The human telomerase catalytic subunit hTERT: organization of the gene and characterization of the promoter. Hum Mol Genet. 1999;8(1):137-142. [Google Scholar] [CrossRef] |
| 10. | Sze KM, Ho DW, Chiu YT, Tsui YM, Chan LK, Lee JM, et al. Hepatitis B Virus-Telomerase Reverse Transcriptase Promoter Integration Harnesses Host ELF4, Resulting in Telomerase Reverse Transcriptase Gene Transcription in Hepatocellular Carcinoma. Hepatology. 2021;73(1):23-40. [Google Scholar] [CrossRef] |
| 11. | Li CL, Hsu CL, Lin YY, Ho MC, Hu RH, Chen CL, et al. HBV DNA Integration into Telomerase or MLL4 Genes and TERT Promoter Point Mutation as Three Independent Signatures in Subgrouping HBV-Related HCC with Distinct Features. Liver Cancer. 2024;13(1):41-55. [Google Scholar] [CrossRef] |
| 12. | Jang JW, Kim HS, Kim JS, Lee SK, Han JW, Sung PS, et al. Distinct Patterns of HBV Integration and TERT Alterations between in Tumor and Non-Tumor Tissue in Patients with Hepatocellular Carcinoma. Int J Mol Sci. 2021;22(13):7056. [Google Scholar] [CrossRef] |
| 13. | Pinyol R, Torrecilla S, Wang H, Montironi C, Pique-Gili M, Torres-Martin M, et al. Molecular characterisation of hepatocellular carcinoma in patients with non-alcoholic steatohepatitis. J Hepatol. 2021;75(4):865-78. [Google Scholar] [CrossRef] |
| 14. | Peneau C, Imbeaud S, La Bella T, Hirsch TZ, Caruso S, Calderaro J, et al. Hepatitis B virus integrations promote local and distant oncogenic driver alterations in hepatocellular carcinoma. Gut. 2022;71(3):616-626. [Google Scholar] [CrossRef] |
| 15. | Li T, Fan J, Wang B, Traugh N, Chen Q, Liu JS, et al. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017;77(21):e108-e110. [Google Scholar] [CrossRef] |
| 16. | Lee JH, Ku JL, Park YJ, Lee KU, Kim WH, Park JG. Establishment and characterization of four human hepatocellular carcinoma cell lines containing hepatitis B virus DNA. World J Gastroenterol. 1999;5(4):289-295. [Google Scholar] [CrossRef] |
| 17. | Luo Y, Hitz BC, Gabdank I, Hilton JA, Kagda MS, Lam B, et al. New developments on the Encyclopedia of DNA Elements (ENCODE) data portal. Nucleic Acids Res. 2020;48(D1):D882-D889. [Google Scholar] [CrossRef] |
| 18. | Niinep K, Anier K, Etelainen T, Piepponen P, Kalda A. Repeated Ethanol Exposure Alters DNA Methylation Status and Dynorphin/Kappa-Opioid Receptor Expression in Nucleus Accumbens of Alcohol-Preferring AA Rats. Front Genet. 2021;12:750142. [Google Scholar] [CrossRef] |
| 19. | Tornesello ML, Cerasuolo A, Starita N, Amiranda S, Bonelli P, Tuccillo FM, et al. Reactivation of telomerase reverse transcriptase expression in cancer: the role of TERT promoter mutations. Front Cell Dev Biol. 2023;11:1286683. [Google Scholar] [CrossRef] |
| 20. | Penev A, Markiewicz-Potoczny M, Sfeir A, Lazzerini Denchi E. Stem cells at odds with telomere maintenance and protection. Trends Cell Biol. 2022;32(6):527-536. [Google Scholar] [CrossRef] |
| 21. | Muller M, Bird TG, Nault JC. The landscape of gene mutations in cirrhosis and hepatocellular carcinoma. J Hepatol. 2020;72(5):990-1002. [Google Scholar] [CrossRef] |
| 22. | Horn S, Figl A, Rachakonda PS, Fischer C, Sucker A, Gast A, et al. TERT promoter mutations in familial and sporadic melanoma. Science. 2013;339(6122):959-961. [Google Scholar] [CrossRef] |
| 23. | Akuta N, Kawamura Y, Kobayashi M, Arase Y, Saitoh S, Fujiyama S, et al. TERT Promoter Mutation in Serum Cell-Free DNA Is a Diagnostic Marker of Primary Hepatocellular Carcinoma in Patients with Nonalcoholic Fatty Liver Disease. Oncology. 2021;99(2):114-123. [Google Scholar] [CrossRef] |
| 24. | Akuta N, Kawamura Y, Suzuki F, Kobayashi M, Arase Y, Saitoh S, et al. Serum TERT C228T is an important predictor of non-viral liver cancer with fatty liver disease. Hepatol Int. 2022;16(2):412-422. [Google Scholar] [CrossRef] |
| 25. | Jung P, Menssen A, Mayr D, Hermeking H. AP4 encodes a c-MYC-inducible repressor of p21. Proc Natl Acad Sci U S A. 2008;105(39):15046-15051. [Google Scholar] [CrossRef] |
| 26. | Jung P, Hermeking H. The c-MYC-AP4-p21 cascade. Cell Cycle. 2009;8(7):982-989. [Google Scholar] [CrossRef] |
| 27. | Jackstadt R, Roh S, Neumann J, Jung P, Hoffmann R, Horst D, et al. AP4 is a mediator of epithelial-mesenchymal transition and metastasis in colorectal cancer. J Exp Med. 2013;210(7):1331-1350. [Google Scholar] [CrossRef] |
| 28. | Xue C, Yu DM, Gherardi S, Koach J, Milazzo G, Gamble L, et al. MYCN promotes neuroblastoma malignancy by establishing a regulatory circuit with transcription factor AP4. Oncotarget. 2016;7(34):54937-54951. [Google Scholar] [CrossRef] |
| 29. | Boboila S, Lopez G, Yu J, Banerjee D, Kadenhe-Chiweshe A, Connolly EP, et al. Transcription factor activating protein 4 is synthetically lethal and a master regulator of MYCN-amplified neuroblastoma. Oncogene. 2018;37(40):5451-5465. [Google Scholar] [CrossRef] |
| 30. | Song J, Xie C, Jiang L, Wu G, Zhu J, Zhang S, et al. Transcription factor AP-4 promotes tumorigenic capability and activates the Wnt/beta-catenin pathway in hepatocellular carcinoma. Theranostics. 2018;8(13):3571-3583. [Google Scholar] [CrossRef] |
| 31. | Huang T, Chen QF, Chang BY, Shen LJ, Li W, Wu PH, et al. TFAP4 Promotes Hepatocellular Carcinoma Invasion and Metastasis via Activating the PI3K/AKT Signaling Pathway. Dis Markers. 2019;2019:7129214. [Google Scholar] [CrossRef] |
| 32. | Papathanasiou M, Tsiftsoglou SA, Polyzos AP, Papadopoulou D, Valakos D, Klagkou E, et al. Identification of a dynamic gene regulatory network required for pluripotency factor-induced reprogramming of mouse fibroblasts and hepatocytes. EMBO J. 2021;40(1):e102236. [Google Scholar] [CrossRef] |
| 33. | Lobanenkov VV, Nicolas RH, Adler VV, Paterson H, Klenova EM, Polotskaja AV, et al. A novel sequence-specific DNA binding protein which interacts with three regularly spaced direct repeats of the CCCTC-motif in the 5'-flanking sequence of the chicken c-myc gene. Oncogene. 1990;5(12):1743-1753. [Google Scholar] |
| 34. | Hark AT, Schoenherr CJ, Katz DJ, Ingram RS, Levorse JM, Tilghman SM. CTCF mediates methylation-sensitive enhancer-blocking activity at the H19/Igf2 locus. Nature. 2000;405(6785):486-489. [Google Scholar] [CrossRef] |
| 35. | Chao W, Huynh KD, Spencer RJ, Davidow LS, Lee JT. CTCF, a candidate trans-acting factor for X-inactivation choice. Science. 2002;295(5553):345-347. [Google Scholar] [CrossRef] |
| 36. | Renaud S, Loukinov D, Bosman FT, Lobanenkov V, Benhattar J. CTCF binds the proximal exonic region of hTERT and inhibits its transcription. Nucleic Acids Res. 2005;33(21):6850-6860. [Google Scholar] [CrossRef] |
| 37. | Chen L, Zhu ZZ, Liu SF, Wan X, Wen BJ, Jiang H, et al. Loss at 16q22.1 identified as a risk factor for intrahepatic recurrence in hepatocellular carcinoma and screening of differentially expressed genes. Neoplasma. 2016;63(1):114-120. [Google Scholar] [CrossRef] |
| 38. | Zhang B, Zhang Y, Zou X, Chan AW, Zhang R, Lee TK, et al. The CCCTC-binding factor (CTCF)-forkhead box protein M1 axis regulates tumour growth and metastasis in hepatocellular carcinoma. J Pathol. 2017;243(4):418-430. [Google Scholar] [CrossRef] |
| 39. | Sun L, Huang C, Zhu M, Guo S, Gao Q, Wang Q, et al. Gastric cancer mesenchymal stem cells regulate PD-L1-CTCF enhancing cancer stem cell-like properties and tumorigenesis. Theranostics. 2020;10(26):11950-11962. [Google Scholar] [CrossRef] |
| 40. | Eldholm V, Haugen A, Zienolddiny S. CTCF mediates the TERT enhancer-promoter interactions in lung cancer cells: identification of a novel enhancer region involved in the regulation of TERT gene. Int J Cancer. 2014;134(10):2305-2313. [Google Scholar] [CrossRef] |
| 41. | Jiang J, Zhao LJ, Zhao C, Zhang G, Zhao Y, Li JR, et al. Hypomethylated CpG around the transcription start site enables TERT expression and HPV16 E6 regulates TERT methylation in cervical cancer cells. Gynecol Oncol. 2012;124(3):534-541. [Google Scholar] [CrossRef] |
| 42. | Deng J, Zhou D, Zhang J, Chen Y, Wang C, Liu Y, et al. Aberrant methylation of the TERT promoter in esophageal squamous cell carcinoma. Cancer Genet. 2015;208(12):602-609. [Google Scholar] [CrossRef] |
| 43. | Lee DD, Komosa M, Nunes NM, Tabori U. DNA methylation of the TERT promoter and its impact on human cancer. Curr Opin Genet Dev. 2020;60:17-24. [Google Scholar] [CrossRef] |
| 44. | Medici V, Schroeder DI, Woods R, LaSalle JM, Geng Y, Shibata NM, et al. Methylation and gene expression responses to ethanol feeding and betaine supplementation in the cystathionine beta synthase-deficient mouse. Alcohol Clin Exp Res. 2014;38(6):1540-1549. [Google Scholar] [CrossRef] |
| 45. | Zhang Y, Yuan T, Su Z, Wang X, Wang Y, Ni Y, et al. Reduced methylation of PP2Ac promotes ethanol-induced lipid accumulation through FOXO1 phosphorylation in vitro and in vivo. Toxicol Lett. 2020;331:65-74. [Google Scholar] [CrossRef] |
| 46. | Pal-Bhadra M, Bhadra U, Jackson DE, Mamatha L, Park PH, Shukla SD. Distinct methylation patterns in histone H3 at Lys-4 and Lys-9 correlate with up- & down-regulation of genes by ethanol in hepatocytes. Life Sci. 2007;81(12):979-987. [Google Scholar] [CrossRef] |
| 47. | Sze KM, Chu GK, Lee JM, Ng IO. C-terminal truncated hepatitis B virus x protein is associated with metastasis and enhances invasiveness by C-Jun/matrix metalloproteinase protein 10 activation in hepatocellular carcinoma. Hepatology. 2013;57(1):131-139. [Google Scholar] [CrossRef] |
| 48. | O'Callaghan NJ, Fenech M. A quantitative PCR method for measuring absolute telomere length. Biol Proced Online. 2011;13:3. [Google Scholar] [CrossRef] |

![]()
Copyright © 2026 Pivot Science Publications Corp. - unless otherwise stated | Terms and Conditions | Privacy Policy




