
Cancer Heterogeneity and Plasticity ISSN 2818-7792
Cancer Heterogeneity and Plasticity 2024;1(2):0007 | https://doi.org/10.47248/chp2401020007
Review Open Access
Function of noncoding RNA in regulating cancer cell plasticity
Peter Hyunwuk Her
1,2,†

 ,
Magnus Lam
1,2,†
,
Sarah Zeng
2
,
Housheng Hansen He
1,2
,
Magnus Lam
1,2,†
,
Sarah Zeng
2
,
Housheng Hansen He
1,2
Correspondence: Housheng Hansen He
Academic Editor(s): Dean G. Tang
Received: Aug 28, 2024 | Accepted: Oct 22, 2024 | Published: Nov 12, 2024
© 2024 by the author(s). This is an Open Access article distributed under the terms of the Creative Commons License Attribution 4.0 International (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium or format, provided the original work is correctly credited.
Cite this article: Her P, Lam M, Zeng S, He H. Function of noncoding RNA in regulating cancer cell plasticity. Cancer Heterog Plast. 2024;1(2):0007. https://doi.org/10.47248/chp2401020007
Recent advances have brought non-coding RNAs (ncRNAs) into the spotlight, revealing their critical regulatory roles in cancer cell plasticity. ncRNAs, such as microRNAs (miRNAs), transfer RNAs (tRNAs), long non-coding RNAs (lncRNAs) and circular RNAs (circRNAs), are now recognized as key players in cellular processes such as chromatin remodeling, mRNA stability, and translation. This review delves into the diverse functions of ncRNAs in stem cells and cancer stem cells (CSCs) biology, emphasizing their impact on maintaining and modulating cellular states. We explore the mechanisms by which ncRNAs influence stem cell self-renewal and differentiation, including their roles in establishing pluripotency and directing differentiation. In the context of cancer, ncRNAs are pivotal in driving processes like epithelial-mesenchymal transition (EMT), which underlies metastasis and therapy resistance. By regulating gene expression and epigenetic landscapes, ncRNAs sustain the dynamic nature of CSCs, facilitating tumor growth and heterogeneity. The review also highlights the potential clinical applications of ncRNAs as biomarkers and therapeutic targets. Advances in ncRNA detection and manipulation have opened new avenues for developing diagnostic tools and innovative treatments. Liquid biopsies, which utilize ncRNAs from biological fluids, provide a minimally invasive approach to monitor tumor dynamics and progression. Uncovering the intricate networks regulated by ncRNAs makes it evident that these molecules play central roles in understanding cancer cell plasticity. Insights into their functions offer promising strategies for targeted cancer therapies, aiming to disrupt the adaptability of cancer cells and improve treatment outcomes.
Keywordsnoncoding RNA, lineage plasticity, cancer stem cell, biomarker, therapeutic target
It is estimated that up to 90% of the genome is actively transcribed into RNAs, yet only 1.5%–2.0% of the human genome consists of protein-coding genes [1–3]. In recent years, it has become clear that the non–protein-coding portion of the genome is not merely spurious transcriptional noise, as previously thought. Particularly, recent deep sequencing has revealed thousands of non-coding RNAs (ncRNAs) with diverse functions in gene expression and genomic architecture [4–7]. ncRNAs are a diverse class of RNA molecules that do not encode proteins but have regulatory roles in gene expression and cellular processes from chromatin remodeling to mRNA stability and translation [8]. ncRNAs can be classified into housekeeping and regulatory ncRNAs with two major size differences, short and long, based on the 200-nucleotide cutoff in mature transcript length [8,9]. Housekeeping ncRNAs, as their name suggests, are critical for general cellular function and demonstrate abundance in the cell. For example, circHIPK3, a circularized splicing product of HIPK3 precursor RNA, is essential for sponging of multiple miRNAs and modulating cell growth [10,11]. Some tRNA genes in the human genome, such as n-Tr20, have also been shown to regulate mTORC1 signaling and consequently reprogramming the synaptic transmission pathways of neuronal cells [12]. However, there is growing evidence of housekeeping ncRNAs contributing to cancer pathogenesis as well [13–15]. Short ncRNAs encompass well-characterized species such as microRNAs (miRNAs), small interfering RNAs (siRNAs), small nuclear RNAs, ribosomal RNAs, and transfer RNAs (tRNAs). Long noncoding RNAs (lncRNAs) can be categorized into various subgroups based on their genomic location and evolutionary origin, including long intergenic RNAs (lincRNAs), antisense RNAs, sense intronic RNAs, enhancer RNAs (eRNAs), circular RNAs (circRNAs) and pseudogenes. Extensive reviews have examined and summarized the oncogenic and tumor suppressive functions of noncoding transcripts, which are particularly evident for miRNAs [16–19], lncRNAs [20–23], and circRNAs [24,25].
Stem cells are unique in their ability to self-renew and differentiate into various cell types. Plasticity is a key feature of stem cells, regulated by reversible epigenetic modifications. While gene-restriction programs are set during embryonic development as cell lineages form, stem cells maintain a level of flexibility for tissue regeneration [24–27]. In cancer, normal stem cell processes are often hijacked to confer malignant properties. Previous studies have identified the presence of cancer stem cells (CSCs) in various solid tumors such as breast, brain, melanoma, pancreatic, prostate, and ovarian cancers [28–34]. These CSCs exhibit properties akin to normal stem cells, including self-renewal, quiescence, and the ability to generate heterogeneous tumor cell populations. They also exhibit resistance to chemotherapy and radiotherapy [28]. This plasticity is largely governed by complex networks of genetic and epigenetic factors. More recently, ncRNAs have been implicated in maintaining cellular plasticity for both stem cells and CSCs. In normal stem cell fate, ncRNAs are crucial for establishing the pluripotent network and are vital for reprogramming somatic cells to achieve pluripotency [35]. Their involvement in cancer cell plasticity is becoming increasingly recognized, particularly in the context of epithelial-mesenchymal transition (EMT), a key process in cancer metastasis that contributes to therapy resistance [36]. In addition, ncRNAs are gaining increasing attention for their large number, specific expression profiles, functional roles, and potential clinical applications.
This review explores the role of ncRNAs in cellular plasticity in more recent advancements, with a focus on miRNAs, lncRNAs, circRNAs and tRNAs in stem cells, CSCs and subsequently the heterogeneity that exists in solid tumors, highlighting their functional relevance in maintaining and modulating cellular states. Potential therapeutic targets and strategies towards ncRNAs will also be explored.
Small ncRNAs, characterized by their length of 20–30 nucleotides and their association with Argonaute family proteins, are categorized into three classes in animals: miRNAs, siRNAs, and PIWI-interacting RNAs [37]. Among these, miRNAs are the predominant class of small RNAs in most somatic tissues. Over 60% of human protein-coding genes have at least one conserved miRNA-binding site. When including the numerous non-conserved sites, it is likely that most protein-coding genes are regulated by miRNAs. miRNAs are small RNAs (approximately 22 nucleotides) that regulate gene expression post-transcriptionally by binding to the 3' untranslated regions (UTR) of target mRNAs, leading to their degradation or translational repression [37]. The biogenesis of miRNAs begins in the nucleus, where primary miRNAs are transcribed by RNA polymerase II, then processed by the Drosha-DGCR8 complex into precursor miRNAs, which are then exported to the cytoplasm. In the cytoplasm, Dicer further processes precursor miRNAs into mature miRNAs, which are incorporated into the RNA-induced silencing complex (RISC) to regulate target mRNAs [38,39].
The most recent version of the miRBase database (v22) includes 38,589 entries representing hairpin precursor microRNAs from 271 organisms, though the functional relevance of many remains to be elucidated [40]. The dysregulation of miRNAs has been associated with cancer development and progression in various cancers [38]. For example, in pancreatic cancer with mutant KRAS, RAS-responsive element-binding protein 1 (RREB1) represses the promoters of miR-143 and miR-145. Simultaneously, both KRAS and RREB1 are targets of miR-143 and miR-145, creating a feedforward mechanism that amplifies the effect of RAS signaling [41]. The clinical utility of miRNAs is highlighted by aberrant miRNA levels indicating the physiological state of cancer cells, which can be detected through miRNA expression profiling and utilized for diagnosis and prognosis. In fact, miRNA profiling is often more accurate than mRNA profiling for classifying tumors because miRNA expression closely correlates with tumor origin and stage. It can also effectively classify poorly differentiated tumors, which are challenging to identify using standard histological methods [42–45]. Furthermore, circulating miRNAs in serum may serve as diagnostic tools. For instance, miR-141 expression levels in serum were reported to be elevated in patients with prostate cancer compared to healthy control individuals [46,47].
Over the past decade, research has increasingly shown that the genomes of various species are extensively transcribed, leading to the production of numerous lncRNAs. lncRNAs are longer than 200 nucleotides and participate in a wide range of regulatory functions, including chromatin remodeling, transcriptional regulation, and post-transcriptional processing [48–50]. This broad definition includes a vast and highly diverse collection of intergenic transcripts, eRNAs, and sense or antisense transcripts overlapping other genes that vary in their biogenesis and genomic origin [51]. Most lncRNAs are transcribed by RNA polymerase II and undergo similar processing steps as mRNAs, including capping, splicing, and polyadenylation. However, lncRNAs typically do not encode proteins. Instead, they function through various mechanisms, including acting as molecular scaffolds, guides, decoys, and enhancers [49]. The resulting lncRNAs are typically capped with 7-methylguanosine at their 5′ ends and polyadenylated at their 3′ ends [52].
Data from the Human GENCODE project indicate that the human genome contains over 16,000 lncRNA genes; however, other estimates suggest there could be more than 100,000 human lncRNAs [53,54]. These lncRNAs are implicated in acquiring all the hallmarks of cancer, ranging from intrinsic abilities like enhanced proliferation and survival to changes in metabolism and interactions with the tumor microenvironment (TME). They are transcriptionally regulated by key oncogenic or tumor-suppressive transcription factors, including p53 [55,56], MYC [57,58], the estrogen receptor [59], and signaling pathways such as Notch [60]. This regulation allows lncRNAs to contribute significantly to oncogenic or tumor-suppressive responses. Their tissue- and condition-specific expression patterns suggest strong potential as biomarkers as well, making them promising targets for clinical interventions. One such lncRNA, the tissue differentiation-inducing non-protein coding RNA (TINCR), is notably dysregulated in various cancers and influences tumor development and progression [61].
circRNAs are produced through an inefficient back-splicing process, in which a downstream splice donor is joined to an upstream splice acceptor, thus creating a covalently closed looped structure that lacks poly-adenylated tails. This process largely utilizes the canonical RNA splicing machinery. Most current research has focused on circRNAs derived from known genes, where in some cases, the circular isoform is much more predominant than its linear counterpart [10]. While escaping debranching can lead to the formation of intronic circRNAs, different mechanisms are involved in the generation of circRNAs comprising exonic sequences [62]. Various regulatory sequences [63,64] and protein factors [65–68] are implicated in this process, with circRNAs being produced both co-transcriptionally and post-transcriptionally [69,70]. circRNAs can act as miRNA sponges, regulate transcription, and interact with RNA-binding proteins.
The number of unique circRNAs produced in human cells (approximately 100,000) [71] is almost fivefold higher than that of protein-coding genes (approximately 20,000) [71–74]. Despite their widespread presence, most circRNAs have not been functionally characterized, and their biological roles remain largely unknown. However, their clinical utility is highlighted through their association with various physiological conditions and cellular features as well as with clinical characteristics such as histological grade, tumor size, metastasis stage, and cancer aggressiveness [75,76]. For example, CiRS-7/CDR1as is highly abundant in intratumoral stromal cells, which are widely used as a prognostic factor in colon and breast carcinomas [77,78]. Furthermore, the tissue and cell-type specificity enables better correlation with specific pathologies, allowing for improved differentiation between cancer subtypes [79–82].
tRNAs are small ncRNA molecules ranging from 56 to 80 nucleotides long and represent a unique clover-leaf secondary structure. The human genome contains genes encoding separate fractions of tRNA species that function either in the cytoplasm or mitochondria (mt-tRNAs) [83–85]. The complex secondary structure of tRNAs consists of 4 major regions: the amino-acid acceptor stem arm, D arm, T arm, and anticodon [86,87]. The anticodon is a triplicate of nucleotide bases which dictate one of the 20 amino acids to be deposited on the 3’ end of the acceptor stem based on the degenerate coding system. Iso-acceptors, therefore, refer to tRNAs of different sequences, which “accept” the same amino acid during the process of mRNA translation.
The unique structural complexity of this small ncRNA population also translates to a wide array of chemical base modifications that each tRNA may harbor, ranging from as many as 14 to 30 modifications [88,89]. Canonically, tRNAs are critical for protein synthesis and draw the bridging gap from transcriptome to proteome translation, where the degenerate coding system is thought to have been fully explored. However, recent studies have also revealed the cleavage of tRNA species by angiogenin nucleases into smaller fragments – tRFs. tRFs expose sequence motifs that would otherwise be hidden in the uncleaved and folded native structure, and contribute to oncogenesis [90–92].
Stem cells are unspecialized cells capable of self-renewal and differentiation into any cell type of the organism, classified based on their developmental potency. Totipotent stem cells, like zygotes, can differentiate into all cell types, including extraembryonic structures [93]. Pluripotent stem cells, such as embryonic stem cells (ESCs) and induced pluripotent stem cells, can form cells of all germ layers but not extraembryonic structures. Multipotent stem cells, like hematopoietic stem cells (HSCs), can specialize in specific cell lineages, while oligopotent and unipotent stem cells have even more restricted differentiation abilities. ESCs, derived from the inner cell mass of the blastocyst, are pluripotent and capable of differentiating into any cell type, though their use in research is ethically debated. Somatic or adult stem cells, found throughout the body, aid in healing, growth, and cell replacement, with mesenchymal stem cells, neural stem cells (NSCs), HSCs, and skin stem cells being notable examples. Below, we summarize the contributions of ncRNAs to each of the three major categories of stem cells.
miRNA profiling during human development reveals specific miRNA signatures at various stages, suggesting roles in early lineage differentiation and developmental processes. These small regulatory RNAs play various roles in cancer. miRNAs influence stem cell behavior through several mechanisms (Figure 1A). They regulate the expression of key transcription factors and signaling pathways that govern stem cell maintenance, differentiation, and self-renewal [94,95]. Additionally, miRNAs can modulate the cell cycle, promoting or inhibiting cell division as needed to maintain stem cell populations. They also participate in fine-tuning cellular responses to environmental cues, such as growth factors and cytokines, which further influences stem cell fate decisions. Furthermore, miRNAs are involved in epigenetic regulation by interacting with chromatin-modifying enzymes, thereby affecting the transcriptional landscape of stem cells [96]. Collectively, these mechanisms highlight the pivotal role of miRNAs, explained in further detail below.

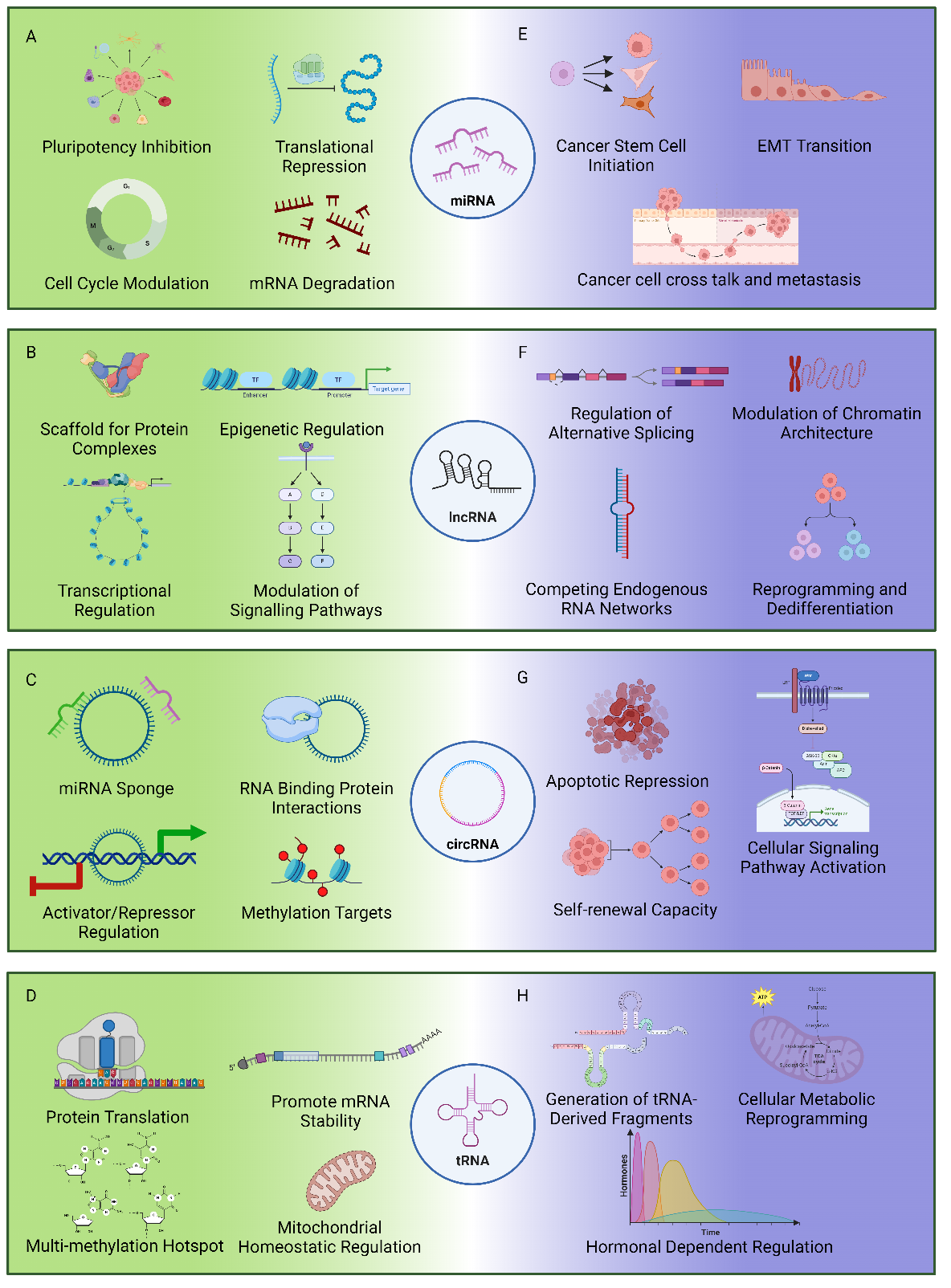
Figure 1. Roles of ncRNAs in Stem Cells (A to D) and CSCs (E to H). (A) miRNAs in stem cells. miRNAs play multiple roles in balancing self-renewal and differentiation. They achieve this balance through translational repression and mRNA degradation, controlling the levels of proteins vital for stem cell maintenance and lineage specification. Additionally, miRNAs modulate the cell cycle, ensuring proper progression through different phases essential for self-renewal and differentiation. (B) lncRNAs in stem cells. lncRNAs serve as scaffolds for protein complexes, facilitating the assembly of multi-protein complexes that are essential for various cellular functions. They also play a significant role in epigenetic regulation by interacting with chromatin modifiers to influence gene expression. Additionally, lncRNAs are involved in transcriptional regulation, where they modulate the activity of transcription factors and RNA polymerase. Furthermore, lncRNAs modulate signaling pathways, acting as key regulators of cellular signaling networks. (C) circRNAs in stem cells. circRNAs act as miRNA sponges, sequestering microRNAs and preventing them from binding to their target mRNAs, thereby regulating gene expression. They interact with RNA-binding proteins, influencing the stability, localization, and translation of mRNAs. Additionally, circRNAs are involved in activator/repressor regulation, modulating transcriptional activities. They also serve as targets for methylation, affecting gene expression and cellular functions. (D) tRNAs in stem cells. tRNAs play a fundamental role in protein translation, facilitating the translation of mRNA into proteins. They promote mRNA stability by protecting mRNAs from degradation, thereby ensuring efficient protein synthesis. tRNAs also serve as multi-methylation hotspots, influencing gene expression and cellular functions through epigenetic modifications. Additionally, tRNAs are involved in mitochondrial homeostatic regulation, maintaining mitochondrial function and energy production. (E) miRNAs in cancer cell plasticity. miRNAs facilitate CSC initiation and maintenance, which ultimately drive tumor initiation, growth, and recurrence. They also regulate EMT, enabling cancer cells to gain migratory and invasive properties necessary for metastasis. Moreover, miRNAs promote cancer cell crosstalk and metastasis by facilitating communication between cancer cells and their microenvironment, aiding in metastatic spread and colonization of new tissues. (F) lncRNAs in cancer cell plasticity. lncRNAs regulate alternative splicing, influencing the generation of diverse protein isoforms that contribute to cancer progression. They modulate chromatin architecture, affecting the accessibility of DNA to the transcriptional machinery and thus altering gene expression profiles. lncRNAs also participate in ceRNA networks, where they act as sponges for microRNAs, thereby regulating the availability of microRNAs to their target mRNAs. lncRNAs play a crucial role in reprogramming and dedifferentiation, processes that enable cancer cells to acquire stem cell-like properties and contribute to tumor heterogeneity and adaptability. (G) circRNAs in cancer cell plasticity. circRNAs contribute to apoptotic repression, enabling cancer cells to evade programmed cell death. They enhance the self-renewal capacity of CSCs, promoting tumor growth and recurrence. Additionally, circRNAs are involved in activating cellular signaling pathways that drive cancer progression and metastasis. (H) tRNAs in cancer cell plasticity. tRNAs contribute to the generation of tRFs, which can regulate gene expression and cellular processes. They play a role in cellular metabolic reprogramming, allowing cancer cells to adapt their metabolism to support rapid growth and survival. tRNAs also participate in hormone-dependent regulation, influencing hormone levels and signaling pathways that are crucial for cancer progression.
Multiple layers of control regulate pluripotency and differentiation in human ESCs (hESCs). At the transcriptional level, the core factors OCT4, NANOG, and SOX2 establish a positive autoregulatory loop crucial for maintaining an undifferentiated state. On the post-transcriptional level, these transcription factors are directly repressed by miR-145 through targeting of their 3’ UTR, thereby controlling the rate of differentiation of hESCs away from their pluripotent state [97,98]. Additionally, recent findings reveal that OCT4 and miR-302 collaborate to repress NR2F2 at the transcriptional and post-transcriptional levels in undifferentiated hESCs. During differentiation, the decline of OCT4 and miR-302 permits the activation of NR2F2, which in turn inhibits OCT4, creating a feedback loop. This intricate interplay of transcriptional and post-transcriptional mechanisms not only sustains the pluripotency of hESCs but also facilitates the precise specification of neuroectoderm lineages during differentiation [99].
HSCs are affected by aging, leading to a decline in their self-renewal and regeneration capabilities, independent of their microenvironment. miRNAs play crucial roles in maintaining HSC function and can modulate tissue senescence in specific cell types [100,101]. miR-146a has been found to regulate the inflammatory response and maintain HSC homeostasis. Depletion of miR-146a leads to the fatigue of HSCs, the development of hematological tumors, and myeloproliferative neoplasms. It has been linked to chronic inflammation and age-related decline in HSC activity [102]. Another important miRNA, let-7, is highly active during embryogenesis and brain development and continues to be expressed in adult tissues [103,104]. In addition to its role in development, let-7 plays a critical function in regulating HSC fate, working alongside miR-99a/100, miR-125b-1/2, and LIN28B to control processes such as self-renewal, proliferation, quiescence, and differentiation [104,105]. Cluster 1-a (let-7a-2, miR-100, miR-125b-1) and Cluster 1-b (let-7c, miR-99a, miR-125b-2) are involved in maintaining HSPC homeostasis, primarily by inhibiting the TGFβ pathway and enhancing Wnt signaling [106]. Conversely, LIN28B represses let-7, inhibiting erythroid development and maintaining stemness.
MicroRNAs regulate neurogenesis through a feedback mechanism involving transcription factors and epigenetic factors. miR-9 and the nuclear receptor TLX form a feedback loop that regulates neural stem cell proliferation and differentiation. miR-9 targets TLX, reducing its expression to promote differentiation, while TLX represses miR-9 to maintain stem cell self-renewal [107,108].
Recent literature suggests that close to a thousand lncRNAs have been identified as potential regulators of stem cell renewal [109]. lncRNAs influence stem cell behavior through several key mechanisms (Figure 1B). They act as molecular scaffolds, bringing together protein complexes and directing them to specific genomic locations, thereby modulating gene expression crucial for stem cell maintenance and differentiation. Additionally, lncRNAs can function as decoys, sequestering transcription factors, miRNAs, or other regulatory molecules away from their targets, thus impacting the transcriptional and post-transcriptional landscape of stem cells. They also play roles in chromatin remodeling by interacting with chromatin-modifying enzymes and altering the epigenetic state of stem cell-specific genes. Moreover, lncRNAs can serve as guides, directing chromatin-modifying complexes to specific genomic loci to activate or repress transcription. They also play a role in regulating post-translational modifications by acting as scaffolds. Several of those roles are highlighted below.
The lncRNA Panct1 is involved in the pluripotency pathway in mouse ESCs. Panct1 interacts with the protein TOBF1, recruiting it to the promoters of pluripotency genes during the early G1 phase of the cell cycle. This recruitment is crucial for the maintenance of the pluripotent state. Loss of Panct1 and TOBF1 leads to the downregulation of pluripotency factors, indicating the essential role of lncRNAs in regulating ESC identity through protein interactions [110]. The lncRNA Xist plays a key role in regulating the fate of ESCs by initiating gene silencing through coating one of the two X chromosomes in female mammals. This process prevents gene dosage imbalance between females and males [111]. Xist further influences ESC development through the use of TALE-based designer transcription factors, guiding cell differentiation and maintaining proper cellular function [112,113].
The lncRNA H19 maintains the quiescent state of long-term hematopoietic stem cells (LT-HSCs), which are responsible for sustaining hematopoiesis over the lifetime of an organism. H19 prevents LT-HSCs from excessive proliferation, which is essential for the long-term maintenance and self-renewal of HSCs [114,115]. It also acts as a molecular sponge for microRNAs that target Igf2, thus modulating the levels of IGF2 protein, which in turn interacts with the IGF1 receptor on HSCs.
Over the years, there has been increasing evidence of lncRNAs contributing to the neuronal structural development in murine models. For instance, Evf2 regulates the transcription of Dlx5 and Dlx6 in the developing mouse forebrain by recruiting DLX and MECP2 transcription factors to regulatory elements in the Dlx5/6 intergenic region [116]. Similarly, Lacuna acts as a negative regulator of neurogenesis by inhibiting the differentiation of NSCs into neurons, while also modulating a neighboring NSC differentiation-related gene Eomes [117]. While the aforementioned lncRNAs have a temporal-specific role during embryonic development, other lncRNAs such as pnky have been identified to function in a broader window across different stages of brain development. Pnky interacts with the splicing regulator PTBP1 to maintain NSCs and balance neurogenesis, with its knockdown leading to enhanced neuronal differentiation [118].
Recent literature suggests that numerous circRNAs have been identified that regulate stem cell behavior [119]. circRNAs influence stem cell fate and function through several key mechanisms (Figure 1C). They can act as molecular sponges, sequestering miRNAs and preventing them from repressing their target mRNAs, which can be important for maintaining the balance of gene expression needed for stem cell maintenance and differentiation. Additionally, circRNAs are involved in regulating transcription by interacting with transcription factors and RNA-binding proteins, modulating the transcriptional landscape of stem cells. They also play roles in the splicing process, influencing the production of different isoforms of proteins essential for stem cell functions. Furthermore, circRNAs are implicated in the regulation of apoptosis and cell cycle progression, which are critical for stem cell proliferation and survival. A few examples are provided below.
The circRNA circBIRC6 functions by sponging microRNAs such as miR-34a and miR-145, which are involved in the down-regulation of genes that maintain pluripotency. By absorbing these microRNAs, circBIRC6 prevents the premature differentiation of ESCs, thereby preserving their ability to self-renew [120]. In contrast to the involvement of specific circRNAs, the overall biogenesis of circular isoforms from the linear counterparts was demonstrated to be controlled by specific proteins such as FUS3 [121]. circRNAs regulated by FUS in mice are similarly regulated in human cells, indicating a conserved mechanism across species.
The circRNA cia-cGAS is significantly upregulated in LT-HSCs. This circRNA inhibits the activation of cGAS, an enzyme that triggers type I interferon production, a process that can lead to HSC failure if overactivated. By suppressing cGAS-induced type I interferon production, cia-cGAS ensures the proper function and longevity of HSCs [122]. Interestingly, as HSCs commit to their differentiated lineage, the abundance and expression patterns of circRNA change with a notable rise in key stages such as enucleation and platelet formation [123,124]. While hematopoietic cell-type-specific circRNA profiles have been identified, there is much left to be elucidated functionally.
Distinct circRNA profiles have also been identified in neural stem cells [125]. The circRNA circHIPK2 is highly expressed at the early stages of NSC differentiation, with its levels decreasing over time. Silencing circHIPK2 significantly promoted the differentiation of NSCs into neurons, as evidenced by increased expression of neuronal markers such as TUJ1, NeuN, and MAP2, without affecting their differentiation into astrocytes. Transplantation of NSCs transduced with circHIPK2 siRNA into stroke-induced mice resulted in improved neuronal plasticity and functional recovery, indicating that circHIPK2 acts as a regulatory molecule guiding NSCs towards a neuronal fate [126]. A recent study demonstrated that silencing of circFAT3 affects the differentiation of neural progenitors and leads to changes in gene expression associated with neural development, including FAT4 and ERBB4 [127].
The structural complexity, diverse sites of modifications, and post-transcriptional modifications of tRNAs present a multitude of factors that contribute to their uniqueness and facilitate biological interplay beyond their traditional role in protein synthesis [128,129]. tRNAs influence stem cell behavior through several key mechanisms (Figure 1D). They play a crucial role in maintaining the cellular protein synthesis machinery, ensuring the production of proteins necessary for stem cell renewal and differentiation. Additionally, tRNAs are involved in regulating gene expression through their interactions with ribosomes and other RNA molecules, impacting the translation efficiency and stability of specific mRNAs. Post-transcriptional modifications of tRNAs, such as methylation and pseudouridylation, can affect their stability and function, thereby influencing stem cell fate decisions. Furthermore, tRNAs can act as signaling molecules, participating in stress response pathways and modulating stem cell survival under adverse conditions. Several examples are detailed below.
The role of 5′-tRFs in stem cell differentiation was investigated in murine ESCs (mESCs), revealing their enrichment in differentiated cells. Krishna et al. identified a specific subset of tRNA loci contributing to 5′-tRFs and suggested an angiogenin-independent processing mechanism [129]. The study demonstrates that 5′-tRFs regulate translation at multiple steps, interact with target mRNAs and ribosomal proteins, and confer functional heterogeneity to ribosomes [130]. Notably, tsGlnCTG interacts with the RNA-binding protein Igf2bp1, destabilizing the Igf2bp1-c-Myc complex and reducing c-Myc mRNA stability, crucial for stem cell maintenance and differentiation [131,132]. Despite modest increases in tRF levels, their specific interactions with target transcripts and proteins are enhanced under differentiation conditions, indicating additional regulatory factors discussed below.
Pseudouridine (Ψ) is a universal modification found in all RNA species, predominantly catalyzed by the human pseudouridine synthase 7 (PUS7) enzyme [133]. PUS7 has been reported to be upregulated in HSCs compared to other pseudouridine synthase family members. Functional knockout of PUS7 in hESCs using CRISPR/Cas9 led to a noticeable increase in overall cell size and non-specific cell fate commitment during the differentiation phase. The absence of pseudouridylation impairs hematopoiesis and renders HSCs functionally defective, ultimately driving disease progression. This study also uncovered a previously unidentified subset of transfer RNA-derived fragments (tRFs) as novel binding targets of PUS7, with Ψ playing a regulatory role in protein synthesis rates and stem cell fate establishment during embryogenesis [134].
Compared to the cytoplasmic tRNA with a consistent high ratio of tRNAs to cytoplasmic mRNAs for highly efficient protein translation, the mitochondrial genome only encodes for 22 mt-tRNAs [83] while harboring the same burden of recognizing all 20 amino acids. Consequently, mutations or modification differences applied to mt-tRNAs elicit more deleterious biological effects due to the insufficient availability of tRNA isoacceptor variants. METTL8, a methyltransferase that specifically deposits the m3C modification on mt-tRNA threonine/serine, was shown to be critical for controlling the differentiation capacity of cortical neural stem cells in a mouse model [135]. The deletion of METTL8 significantly suppresses synthesis of mitochondrial proteins, driving the earlier neuronal differentiation. Interestingly, the knockout effects are rescuable through pharmacological enhancement of the respiratory capability in mitochondria.
Cancers originating in different tissue sites are generally further categorized into molecular subtypes, acknowledging that the lineage plasticity of cells or tumors is intertwined with the transition from subtype one to subtype two. This plasticity contributes to the heterogeneity observed in solid tumors. For instance, the most prevalent and well-studied non-cutaneous cancer, breast cancer, is classified into two major histological subtypes: luminal and basal. These subtypes can be further distinguished into four molecular types: estrogen receptor positive/progesterone receptor positive ER+/PR+, estrogen receptor negative/progesterone receptor negative ER-/PR-, HER2+, and Triple Negative Breast Cancer (TNBC) [136]. High ER and PR expression in luminal breast cancer tumors historically have better prognosis than basal tumors. Consequently, previous studies have shown evidence of ER-low luminal tumors undergoing plasticity to gain characteristics of basal-like tumors through drivers such as SOX10 expression, suggesting this plasticity is largely regulated by changes in transcriptomic dynamics [137,138].
On the other hand, the interactions between surrounding metabolites and the TME also influence cellular plasticity from the context of acquiring energy sources for survival and proliferation. Cancer cells are capable of switching between glycolysis, a glucose-dependent pathway, or the mitochondrial oxidative phosphorylation for metabolic functions. Specifically, the growth of cancer cells in a three-dimensional structure recapitulates metabolic plasticity of the TME, which favors mitochondrial-dependent metabolism with greater glycolytic reserve potential and reduced dependency on glutamine supplementation [139]. In prostate cancer, increased levels of glutamine help drive resistance to radiotherapy. Therefore, metabolites also regulate the survival of CSCs in certain diseases [140].
Recent evidence suggests that miRNAs play a regulatory role in cancer cell plasticity [141], impacting processes such as self-renewal, differentiation, and resistance to therapy [142,143] (Figure 1E). They achieve this by modulating gene expression at the post-transcriptional level, which affects the behavior and characteristics of cancer cells. miRNAs can control EMT, a critical process for cancer metastasis, by targeting specific transcription factors and signaling pathways involved in this transition. Additionally, miRNAs influence the TME by altering the expression of genes involved in cell-cell communication and extracellular matrix remodeling. They can be packaged into exosomes and transported to other cells, thereby affecting gene expression and signaling pathways in recipient cells [144]. This crosstalk between cancer cells and their microenvironment facilitated by miRNAs contributes to the dynamic nature of tumor growth and metastasis. Through these mechanisms, miRNAs that are integral to the regulation of cancer cell plasticity are summarized in Table 1, with selected examples elaborated further below.
Table 1. Function of miRNAs in CSCs and their interaction partners
miRNAs function through complex mechanisms that are highly dependent on the specific cell type and molecular context. In glioma stem cells, miR-34a acts as a tumor suppressor by targeting and inhibiting the expression of oncogenes such as c-Met, Notch-1, and Notch-2 [145]. These are key pathways for maintaining the self-renewal and survival of glioma cells. By downregulating these oncogenes, miR-34a induces cell cycle arrest, prevents cell migration, and promotes differentiation, thereby reducing the malignant potential of glioma stem cells. In prostate cancer, miR-34a functions through a different mechanism by directly targeting CD44, a surface adhesion molecule highly expressed in prostate CSCs, which are responsible for tumor initiation, metastasis, and resistance to therapies [146]. miR-34a binds to the 3'-UTR of CD44 mRNA, leading to its degradation and reduced protein expression. The knockdown of CD44 in prostate CSCs phenocopies the effects of miR-34a overexpression, inhibiting clonogenic expansion and metastasis. In vivo studies have shown that systemic delivery of miR-34a significantly reduces metastasis and extends the survival of mice with prostate cancer xenografts. This context-dependent activity of miR-34a underscores the complexity of its regulatory role across various cancers. Its ability to target distinct oncogenes or tumor suppressor genes in different cancer types reflects its potential as a therapeutic tool. This adaptability suggests that its application could be tailored to address the specific molecular pathways that are dysregulated in each cancer type.
miR-31 plays a role in maintaining mammary stem cells and promoting breast cancer development and progression. miR-31 is significantly enriched in breast cancer stem cells (BCSCs) (CD24+CD90+), showing a fivefold increase compared to CD24–CD90– cells [157]. This miRNA favors the expansion of BCSCs and their tumor-initiating capability. Loss of miR-31 reduces metastasis, evidenced by lower levels of EMT-associated factors and metastasis inhibitors, a shift to a mesenchymal-like phenotype, reduced lung metastasis, and higher metastasis-free survival in PyVT/KO mice. Moreover, the expression of miR-31 is regulated by the NF-κB pathway. miR-31 directly targets Wnt antagonists, including AXIN1, GSK3β, and DKK1, thereby activating the Wnt-β-catenin pathway, which ultimately contributes to tumor progression.
The miR-200 family and miR-205 play a crucial role in regulating EMT by targeting the transcriptional repressors ZEB1 and SIP1 [169,170]. These microRNAs are markedly downregulated in cells undergoing EMT, either due to TGF-β exposure or ectopic expression of the protein tyrosine phosphatase Pez. Enforced expression of miR-200 alone can prevent TGF-β-induced EMT, indicating its critical role in maintaining the epithelial phenotype. Conversely, inhibition of these microRNAs induces EMT, characterized by loss of E-cadherin and gain of mesenchymal markers, along with enhanced cell motility. Moreover, the loss of miR-200 and miR-205 is observed in invasive breast cancer cell lines and regions of metaplastic breast cancer specimens, suggesting that mechanisms involved in the downregulation of these microRNAs indeed contribute to tumor progression and metastasis [171].
lncRNAs play pivotal roles in regulating cancer cell plasticity through several key mechanisms (Figure 1F) [172]. One of their most well-documented and well-established functions is their role in regulating alternative splicing, leading to diverse protein isoforms that enhance cancer cell adaptability [173–175]. lncRNAs also act as molecular scaffolds, organizing chromatin-modifying complexes and transcription factors at specific genomic loci, thereby altering chromatin architecture to activate or repress gene expression [176–178]. This impacts key cellular processes such as proliferation, differentiation, and the maintenance of CSC properties. Additionally, lncRNAs serve as competing endogenous RNAs (ceRNAs) by sequestering miRNAs, preventing them from binding to their target mRNAs. They further facilitate crosstalk, between cancer cells and the TME, influencing stromal and immune cell behavior to create a supportive niche for tumor growth and metastasis. lncRNAs also promote reprogramming and dedifferentiation, which enhance the stem-like state and resilience of cancer cells. These aspects are detailed in Table 2, with additional examples explored further below.
Table 2. Function of lncRNAs in CSCs and their interaction partners.
In prostate stem-like cells, H19 has been shown to enhance stemness through its regulation of key pluripotency transcription factors, namely OCT4 and SOX2 [184]. Cells expressing high levels of H19 display stem-like properties, including increased expression of stem cell surface markers, resistance to cell death, and enhanced sphere-forming capacity. Suppressing H19 with siRNA leads to a decrease in OCT4 and SOX2 expression, which in turn reduces colony-forming potential and other stem cell-like characteristics. Conversely, overexpression of H19 enhances these traits, promoting the maintenance of prostate CSCs. This suggests that in prostate cells, H19 directly influences transcriptional regulators which govern stemness and survival. In contrast, in BCSCs, H19 plays a different regulatory role through an H19/let-7/LIN28 pathway, highlighting the complexity of lncRNA functions. In this system, H19 acts as a ceRNA to sponge the tumor-suppressive miRNA let-7, which leads to the upregulation of LIN28, a pluripotency factor that supports BCSC maintenance. LIN28 further inhibits the maturation of let-7, creating a feedback loop that reinforces high levels of H19 and LIN28, thereby sustaining BCSC properties such as self-renewal and tumorigenicity. This reciprocal negative feedback loop is essential for maintaining BCSCs and promoting their therapy resistance [185]. These examples underscore the context-dependent role of lncRNAs.
The lncRNA MALAT1 plays a functional role in regulating cell cycle progression by modulating the expression of the oncogenic transcription factor B-MYB. MALAT1 is differentially expressed throughout the cell cycle and is essential for both the G1/S and mitotic phases. Depletion of MALAT1 activates p53 and its target genes, leading to cell cycle arrest and reduced cellular proliferation. This cell cycle arrest is particularly noteworthy as it implicates p53 as a major downstream mediator of MALAT1 activity. Furthermore, MALAT1 promotes the proliferation and invasiveness of CSCs in pancreatic and liver cancers by modulating the expression and pre-mRNA processing of key transcription factors involved in mitotic progression [186,199]. MALAT1-depleted cells also exhibit defects in chromosome segregation and spindle assembly, highlighting MALAT1’s role in maintaining genomic stability during mitosis [200].
lncRNA HOTAIR interacts with the Polycomb Repressive Complex 2 (PRC2) to modify histones and silence gene expression, affecting EMT and CSC maintenance [187]. In BCSCs, HOTAIR indirectly suppresses miR-7, which normally inhibits the oncogene SETDB1. This suppression facilitates EMT and the maintenance of the CSC pool by upregulating the STAT3 pathway, leading to increased expression of c-myc, Twist, and miR-9 [188]. Moreover, HOTAIR modulates the expression of HoxD10, which contributes to miR-7 downregulation and further promotes tumor invasiveness.
lncRNA SOX2 Overlapping Transcript (SOX2-OT) regulates the expression of the SOX2 gene, a critical factor for maintaining pluripotency and self-renewal in stem cells. This lncRNA enhances the stem-like properties of cancer cells by upregulating SOX2 expression. SOX2-OT is involved in various cancers, where it promotes tumor growth and CSC maintenance by regulating SOX2 levels. This regulation contributes to the aggressiveness and recurrence of these malignancies [190,191,201].
circRNAs contribute to the plasticity of cancer cells under stress and therapeutic pressures through several mechanisms (Figure 1G) [202]. In cancer cells, circRNAs act as molecular sponges for miRNAs to prevent the repression of oncogenic targets, thus promoting cancer progression and metastasis. Additionally, circRNAs interact with RNA-binding proteins and transcription factors to modulate gene expression and stabilize mRNAs related to cancer, leading to changes in cell cycle progression, apoptosis resistance, and enhanced metastatic potential. This modulation of gene expression primarily supports cell survival and proliferation in cancer. Furthermore, circRNAs in cancer cells facilitate dynamic adaptability by regulating apoptosis and cell cycle pathways, enabling cancer cells to evade death and sustain growth. These circRNAs are detailed in Table 3 and further explained below.
Table 3. Function of circRNAs in CSCs and their interaction partners.
The circRNA circAGFG1 was found to be upregulated in colorectal cancer cell lines and its knockdown leads to a marked reduction in cell proliferation, migration, invasion, and stemness, while enhancing apoptosis [205]. This indicates the crucial role of circAGFG1 in maintaining the CSC properties, including self-renewal and metastatic potential. The mechanism involves circAGFG1 sponging miR-4262 and miR-185-5p, which ordinarily suppress the expression of the transcription factor YY1. By sequestering these miRNAs, circAGFG1 facilitates the upregulation of YY1, which in turn promotes the transcription of CTNNB1. The activation of the Wnt/β-catenin signaling pathway through this circAGFG1/miR-4262/miR-185-5p/YY1/CTNNB1 axis enhances CSC properties and contributes to colorectal cancer progression and metastasis.
The circRNA hsa_circ_002178 is significantly upregulated in breast cancer tissues and cell lines and its high expression correlates with poor clinical outcomes, including low survival rates, larger tumor size, lymph node metastasis, and advanced TNM stage. Mechanistically, hsa_circ_002178 promotes the growth, invasion, and migration of breast cancer cells by modulating the miR-1258/KDM7A axis. Overexpression of hsa_circ_002178 leads to the formation of microspheres, and increased levels of stemness markers (ALDH1, CD44, NANOG, and OCT4), and elevated ALDH1 activity, which are indicative of enhanced CSC properties. This circRNA acts as a sponge for miR-1258, thereby reducing the miRNA's inhibitory effect on KDM7A. KDM7A, in turn, facilitates the stem-like characteristics of breast cancer cells, contributing to tumor growth and metastasis [206].
The circRNA circCTIC1 was found to be highly expressed in colon tumors and colon tumor-initiating cells (TICs), and its knockdown impaired the self-renewal capacity of these cells, whereas overexpression enhanced it. Mechanistically, circCTIC1 interacts with the nuclear remodeling factor (NURF) complex, recruiting it to the c-Myc promoter, thereby facilitating the transcriptional activation of c-Myc, a core transcription factor for stemness. This interaction underscores the role of circCTIC1 as a protein modulator, driving the self-renewal and tumorigenic potential of colon TICs through the NURF-mediated c-Myc pathway [207].
tRNAs in cancer cells can undergo extensive modifications, enhancing their stability and functionality to meet the increased demands for protein synthesis of rapidly proliferating cancer cells (Figure 1H). These modifications can affect the efficiency and accuracy of translation, promoting the production of proteins that support cancer progression and metastasis [212]. Additionally, tRFs have recently emerged as novel regulators in cancer, where they can act as signaling molecules, influencing gene expression, and modulating cellular stress responses. tRNAs and tRFs are often co-opted to support oncogenic processes, including the regulation of apoptosis, cell cycle progression, and metabolic reprogramming, enabling cancer cells to evade cell death, sustain growth, and adapt to hostile microenvironments. Thus, tRNAs in cancer cells play a multifaceted role in maintaining the plasticity and resilience of cancer cells, distinguishing their function from the more static roles they perform in normal cellular contexts. tRNAs affecting cancer cell plasticity are detailed in Table 4, with some examples highlighted below.
Table 4. Function of tRNAs in CSCs and their interaction partners.
tDR-000620 has been identified as significantly downregulated in the CSCs of TNBC. tDR-000620 was found to play a pivotal role in the suppression of CSC traits, including the inhibition of EMT, recurrence, and metastasis. In TNBC, where the CSC population is highly enriched, the downregulation of tDR-000620 correlates with enhanced self-renewal and tumor-initiating capacities, driving aggressive tumor behavior. Moreover, low tDR-000620 expression was shown to be an independent adverse prognostic factor for recurrence-free survival, further linking its role to poor clinical outcomes. Functional analyses suggest that tDR-000620 is involved in immune-response pathways such as cytokine-cytokine receptor interactions and AMPK signaling, which are essential in the regulation of cell proliferation, metabolism, and stress responses [213].
tRF/miR-1280 has emerged as a key regulator of CSC-like properties and metastasis in colorectal cancer (CRC). This 17-base pair fragment was shown to suppress Notch signaling pathways that support the function and maintenance of CSCs, which are crucial for tumor progression and metastasis. tRF/miR-1280 is significantly downregulated in CRC tissues compared to adjacent normal tissues, and its ectopic expression reduces cell proliferation, colony formation, and metastasis, indicating a tumor-suppressive function. Mechanistically, tRF/miR-1280 directly targets the JAG2 ligand, inhibiting the activation of Notch1 and Notch2 receptors, which are essential for sustaining CSC-like traits in CRC. Through the inactivation of Notch signaling, tRF/miR-1280 leads to the downregulation of critical transcription factors like Gata1 and Gata3, and it increases the expression of miR-200b, a key regulator of EMT. This cascade ultimately results in the repression of ZEB1 and SUZ12, key drivers of CSC maintenance and EMT, thus preventing tumor growth and metastasis [215].
Glycine tRNA-derived fragment (Gly-tRF) enhances liver CSC-like properties and promotes the migration of hepatocellular carcinoma (HCC) cells by targeting NDFIP2, a key regulator of ubiquitin ligase function. Gly-tRF is upregulated in HCC tissues and cell lines, where it drives EMT and increases the stemness and migratory capacity of cancer cells. Mechanistically, Gly-tRF downregulates NDFIP2 by binding to its mRNA 3′-UTR, thereby activating the AKT signaling pathway, a pathway known to contribute to tumor progression and metastasis [216].
The role of ncRNAs in cancer cell plasticity and their contributions to cancer development and progression position them as highly valuable biomarkers with diagnostic, prognostic, and therapeutic potential. Notably, ncRNAs, including CSC-derived ncRNAs, can be detected in biological fluids such as blood and serum. They can be found either as circulating tumor nucleic acids or within secreted circulating tumor cells (CTCs) and extracellular vesicles like exosomes [217]. This capability enables liquid biopsies, offering the advantage of detecting and monitoring the molecular identity and evolution of tumors in a minimally invasive manner, thus facilitating precise prognostic and therapeutic interventions. The applicability of detecting ncRNA expression in liquid biopsies has been validated in the diagnosis, prognosis, and recurrence monitoring of several cancers [218–222]. Liquid biopsies also hold potential in overcoming the limitations of tissue biopsies by identifying molecular heterogeneity between primary and metastatic tumor sites and detecting early tumor formation, unresponsiveness to conventional therapies and disease relapse, often associated with CSC occurrence and their molecular dysregulation at the ncRNA level.
Over the past decade, there has been growing interest in targeting CSC-related or even CSC-specific ncRNAs for CSC elimination. Instead of using drugs to target ncRNAs expressed in CSCs, studies are widely exploring the transport of ncRNA analogs or inhibitors into cells using synthetic nucleic acids. However, many challenges remain in selecting the appropriate CSC-related ncRNA target that is essential for CSC survival and function, as well as accurately evaluating how the introduced oligonucleotide will impact a key biological pathway in CSCs.
The ease of detection of miRNAs in blood and CTCs, coupled with resistance to degradation, underscores their clinical potential [47]. Circulating miRNAs have demonstrated diagnostic power in various cancers, especially gastrointestinal malignancies [223]. For instance, isolating miRNAs from CTCs in colorectal and breast cancers ensures specificity by excluding non-cancer-related miRNAs in the bloodstream [224,225]. However, limitations include low disease and tissue specificity and the potential for non-cancer-related miRNA elevation due to conditions such as inflammation [226]. These challenges are less pronounced in exosome-derived miRNAs, which show higher tissue and cancer type specificity [227]. The diagnostic and prognostic value of miRNAs is supported by their ability to regulate multiple downstream gene targets and pathways involved in oncogenesis and CSC stemness. For instance, miR-548c-3p has been identified as a diagnostic and prognostic marker in prostate cancer, as elevated levels of this miRNA are associated with poor patient survival [228,229]. MiR-548c-3p overexpression is predominantly found in the CD133+α2β1Hi CD44+ prostate cancer subpopulation, where it displays stem cell-like characteristics [230,231]. This miRNA is linked to the activation of stem cell-related genes, enhancing self-renewal and conferring radioresistance.
lncRNAs, known for their tissue-specific expression patterns, are valuable biomarkers for cancer diagnosis and prognosis. Their regulatory roles in CSC pathophysiology suggest significant translational impacts in oncogenesis, tumor prognosis, and therapy [232]. lncRNAs can be detected in body fluids and are resistant to RNase degradation, making them suitable for liquid biopsies [23,233]. Due to their easy detectability, lncRNAs can be employed to predict cancer behavior before, during, or after anti-cancer therapies. For example, PCA3 is a non-coding RNA that is overexpressed in prostate cancer tissues compared to normal tissues. The detection of PCA3 in urine has become a non-invasive diagnostic tool, complementing the traditional prostate-specific antigen test. The PCA3 test helps improve diagnostic accuracy by reducing unnecessary biopsies in patients with elevated PSA levels but a lower risk of aggressive prostate cancer, making it a valuable biomarker in clinical settings [234].
The stability, specificity, and detectability of circRNAs in body fluids, including exosomes, make them promising biomarkers for early cancer detection, monitoring progression, and guiding therapy [235–237]. Studies have shown that circRNA levels can be associated with tumor grade, size, and metastasis, making them valuable tools in cancer diagnostics. For instance, circFARSA levels are elevated in the plasma of non-small cell lung cancer patients, correlating with tumor aggressiveness [238]. Additionally, circRNAs found in exosomes offer potential for early disease detection, as they circulate in body fluids and reflect the molecular state of the tissues from which they originated [239,240].
Chemical modifications of tRNAs, such as methylation, play critical roles in maintaining the fidelity and efficiency of translation, which can be altered in cancer cells to support tumor growth and progression [241]. tRFs are novel biomarkers that can be used for diagnosis, prognosis, and classification of tumor subtypes. Specific types include 5′-tRFs that are upregulated in prostate adenocarcinoma (PRAD) and have been shown to distinguish PRAD tumors from normal tissues with high sensitivity and specificity [242]. Furthermore, a tRF score, based on eight 5′-tRFs, is highly predictive of survival in PRAD patients and offers improved prognostic information when combined with the Gleason score. Based on 5′-tRF expression profiles, four distinct molecular subtypes of PRAD have been identified that are each associated with different survival outcomes and clinicopathological features. These subtypes also exhibit distinct genomic landscapes, offering further prognostic insights.
Oncogenic miRNA expression can be effectively inhibited using antisense oligonucleotides (ASOs), which bind to miRNA targets via base-pair complementarity (Figure 2A). The function of ASOs is enhanced by incorporating various chemical configurations such as locked nucleic acids, anti-miRNA oligonucleotides, and antagomirs, which improve their stability and efficacy [243,244]. For instance, an antagomir specifically designed to target and knockdown the oncogene miR-21 has been employed in breast cancer MCF-7 cells. This intervention resulted in a notable reduction of MCF-7 tumor growth in both in vitro and in vivo tumor xenograft models, achieved by inhibiting cell proliferation and inducing apoptosis [245].
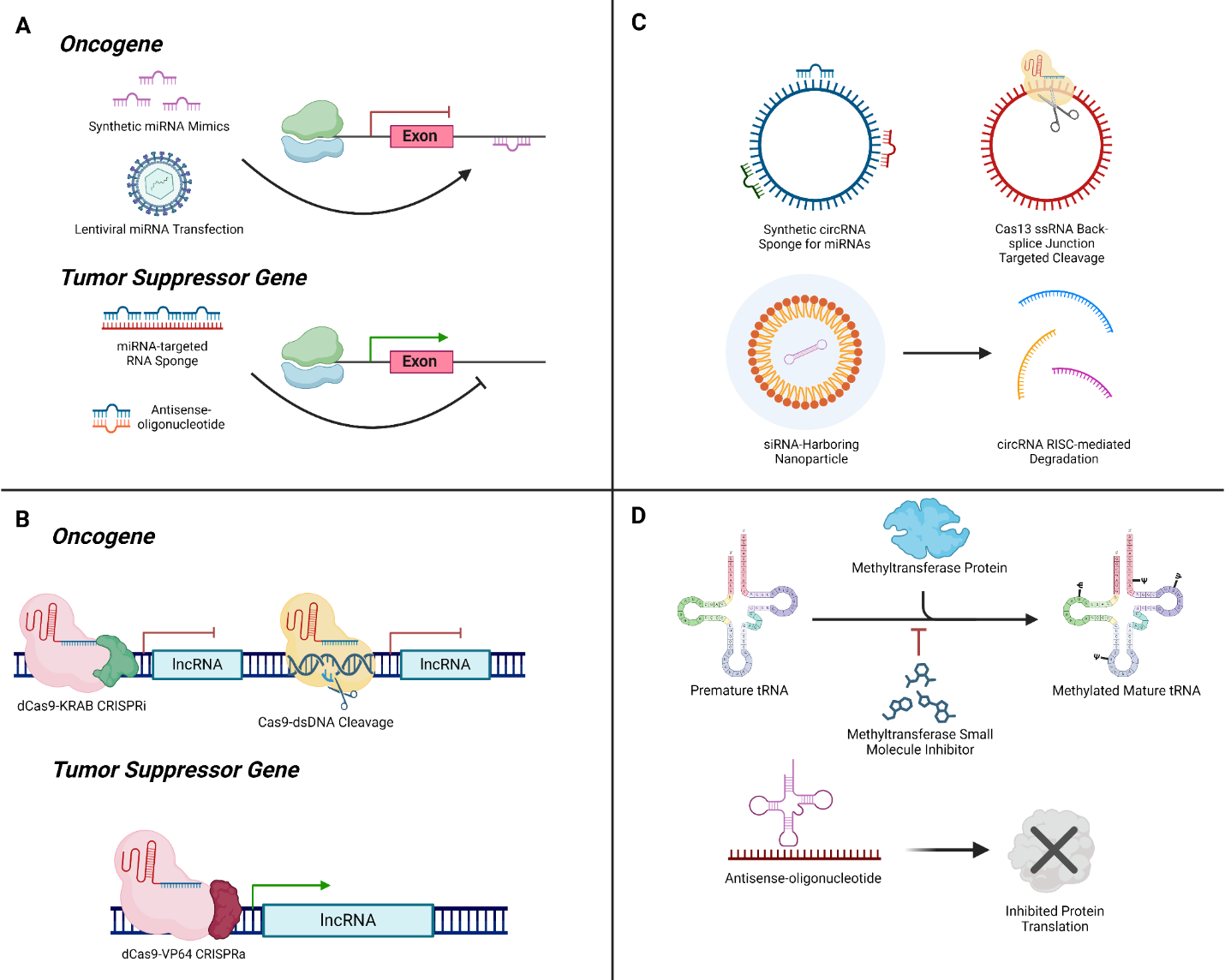
Figure 2. Therapeutic Potentials of ncRNAs. (A) Strategies for targeting oncogenes and tumor suppressor genes using miRNAs. For oncogenes, synthetic miRNA mimics and lentiviral miRNA transfection can inhibit oncogenic pathways. In contrast, for tumor suppressor genes, miRNA-targeted RNA sponges and ASOs can enhance the expression of these genes by sequestering miRNAs that would otherwise repress them. (B) CRISPR-based approaches for regulating oncogenes and tumor suppressor genes through lncRNAs. Oncogenes can be targeted using dCas9-KRAB CRISPR interference (CRISPRi) to inhibit gene expression or Cas9-dsDNA mediated cleavage for gene disruption. Tumor suppressor genes can be activated using dCas9-VP64 CRISPR activation (CRISPRa) to enhance their expression via lncRNAs. (C) Methods for targeting circRNAs in cancer therapy. Synthetic circRNA sponges can sequester miRNAs, while Cas13 ssRNA-mediated back-splice junction targeted cleavage and siRNA-harboring nanoparticles can lead to circRNA degradation through the RISC complex, reducing their oncogenic functions. (D) Approaches for modulating tRNA functions in cancer cells. Inhibition of methyltransferase proteins using small molecule inhibitors can prevent the methylation of tRNAs, thereby disrupting protein translation. Additionally, ASOs can bind to pre-mature tRNAs, preventing their maturation and function in protein synthesis.
Additionally, the suppression of oncogenic miRNAs can be achieved using “miRNA sponges” (Figure 2A). These sponges work by incorporating complementary binding sites for miRNA-targeted RNA into RNA transcripts expressed from strong promoters. This mechanism effectively sequesters miRNAs, preventing them from binding to their endogenous targets, thereby inhibiting miRNA activity in cultured cancer cells and mouse models [243]. Inhibiting miR-22 with a specific “sponge” in LM2 cells, a highly metastatic breast cancer cell line, led to a significant decrease in breast cancer metastasis to the lungs [246]. In contrast, miRNA mimics or lentiviral vectors can be employed to elevate the expression levels of tumor suppressor miRNAs, which are typically found to be underexpressed in cancer cells [247]. RNA interference (RNAi) includes siRNAs and short hairpin RNAs (shRNAs). While siRNAs offer transient gene silencing due to their instability, shRNAs provide durable and long-lasting effects in vivo [248,249]. Virus transfection, the primary method for delivering shRNA plasmids to target sites, can also be employed to introduce exogenously synthesized miRNA plasmids into cancer cells, thereby upregulating the corresponding miRNA.
Additionally, lncRNAs can be targeted therapeutically due to their roles in inducing degradation, regulating transcription, and/or inhibiting interactions with other regulatory factors. Utilizing ASOs to target lncRNAs could be a promising strategy for cancer treatment. For example, the knockdown of MALAT1 with ASOs has been shown to significantly inhibit tumor growth and metastasis in breast cancer [250] and lung cancer [251]. Recent research has demonstrated that CRISPR/Cas9 technology can effectively repress or activate the transcription of loci expressing lncRNAs [252,253] (Figure 2B). The CRISPR/Cas9 system has been utilized to target genomic DNA in cancer cells and animal cancer models. For instance, knocking out the lncRNA NEAT1 and MALAT1 inhibited the metastasis of cancer cells [254,255]. Many lncRNAs are uniquely expressed in various tissues and even among different individuals, making personalized treatment a viable clinical option based on each patient's specific situation. Although CRISPR/Cas9 theoretically offers broad adaptability and target specificity for genome editing, practical applications still face challenges due to potential off-target cleavage events [256,257]. RNAi has also been used to therapeutically target lncRNAs. Adenovirus vectors are widely used, with numerous clinical trials exploring their applications [249,258]. Adeno-associated viruses (AAVs), composed of non-enveloped, single-stranded DNA, are efficient gene delivery systems due to their non-pathogenicity, lack of immune response, and stability in live cells [259]. Optimized AAVs have been developed for targeting lncRNAs in human cancer cells, providing a strong foundation for tumor treatment in clinical settings [237]. Transfecting a gastric cancer cell line with HOTAIR shRNA through retroviral delivery significantly reduced cell spread in peritoneal dissemination [236].
Circular RNA transcripts are currently being developed and evaluated as potential therapeutics both in vitro and in vivo. This includes synthetic circRNAs that can bind to or sponge oncogenic miRNAs [260–262] (Figure 2C). Cancer-promoting circRNAs can be effectively targeted and silenced using RNAi, ASOs, or CRISPR-Cas13 technology by focusing on their unique backsplice junction sequences [263,264]. Nanoparticles have greatly enhanced the practical application of circRNA-based therapeutics in vivo. Nanoparticles are capable of carrying therapeutic agents and precisely delivering them to diseased tissues [265]. Lipid nanoparticles (LNPs) represent the most advanced system for nanoparticle-based drug delivery [266]. Nanoparticle delivery systems effectively address several challenges associated with RNAi molecules by protecting them from degradation, enhancing cellular uptake, and reducing immune activation [267,268]. Gold nanoparticles (AuNPs) conjugated with siRNA targeting circDnmt1 effectively suppressed cellular autophagy, reduced tumor growth, and increased the lifespan of mice [269].
tRNA methyltransferases (TRMs) like hTRM9L and ALKBH8 have been linked to stress responses and cancer phenotypes, suggesting that modulating their activity could suppress cancer cell proliferation while sparing normal cells [269–271]. Furthermore, pseudouridine modifications and the production of tRFs can influence protein synthesis and cell differentiation, highlighting the complexity of tRNA modifications in cancer biology [134]. Understanding these mechanisms opens avenues for developing targeted therapies that disrupt specific tRNA modification pathways, potentially leading to novel cancer treatments that minimize harm to healthy tissues (Figure 2D).
While miRNA-based therapies, including the use of ASOs, miRNA sponges, and miRNA mimics, present promising strategies for modulating gene expression in cancer, potential toxicity remains an undeniable concern. ASOs, particularly those incorporating chemical modifications enhance stability and efficacy but may provoke off-target effects and unintended immune responses [272–274]. Additionally, miRNA sponges and viral vectors used for delivering shRNAs or miRNA mimics may inadvertently interfere with normal cellular functions by sequestering endogenous miRNAs that play critical roles in maintaining homeostasis. Furthermore, prolonged or high-dose administration of these therapeutic agents could disrupt essential pathways, leading to toxicity in non-cancerous tissues [275,276].
While approaches like ASOs and CRISPR/Cas9 offer targeted knockdown or activation of lncRNAs, they may lead to unintended off-target effects, which could disrupt normal cellular processes and induce toxicity. For instance, lncRNAs often play crucial roles in the regulation of multiple biological pathways, and silencing them might interfere with vital functions in non-cancerous tissues. Despite their potential, challenges remain in understanding lncRNAs' cellular functions and their roles in cancer due to their incomplete conservation across species and limited structural and functional data [277,278]. Considering the vast number of deregulated lncRNAs in cancers, there remains a continued need to characterize those that play significant roles in specific processes within particular cancer types, subtypes, or subpopulations, such as CSCs. Additionally, viral vectors used in RNAi-based therapies, such as AAVs, although generally safe, can elicit immune responses or trigger inflammatory pathways, especially when administered at high doses. Furthermore, CRISPR/Cas9 technology carries the risk of off-target cleavage events, which may result in genomic instability or unintended mutations.
While the use of circRNA-based therapeutics and nanoparticles holds great promise, potential toxicity remains a key concern in their clinical application. The long-term biocompatibility and potential adverse effects of these delivery systems must be carefully evaluated. For instance, LNPs, though highly effective in protecting RNA molecules from degradation and enhancing cellular uptake, can trigger immune responses or cause liver toxicity if improperly dosed or formulated. Similarly, while AuNPs have shown potential in targeting specific cancer-related circRNAs, their accumulation in tissues over time may lead to organ toxicity. Challenges in understanding circRNA functionality and molecular targets also persist. Moreover, the lack of discrimination between cancer and non-cancer patients in circulating circRNA levels limits their use as reliable cancer biomarkers. Although the exploration of circRNAs as potential biomarkers in cancer has shown promise, the field is still in its early stages, and much remains to be understood. Currently, there are no clinical trials underway, highlighting the need for further research to fully elucidate their clinical utility.
Toxicity concerns related to therapeutics targeting TRMs stem from the complex role these enzymes play in both cancer and normal cell functions. While modulating TRM activity, such as hTRM9L and ALKBH8, could effectively suppress cancer cell proliferation, there is a risk of off-target effects due to the essential role of tRNA modifications in normal cellular processes. These TRMs are involved in stress responses and protein synthesis regulation in both healthy and cancer cells. Therapies that disrupt specific tRNA modification pathways, such as pseudouridine modifications or tRFs, could inadvertently affect normal cell differentiation and protein production. As a result, there is a fine balance between achieving therapeutic efficacy and minimizing toxicity to healthy tissues. Ensuring selective targeting and reducing adverse side effects will be crucial in developing these therapies for clinical use. Similar to circRNAs, the global role of tRNAs is critical, as they are fundamental to the central dogma of molecular biology by facilitating the translation of RNA into proteins. However, whether targeting tRNAs as a therapeutic strategy is feasible remains open, warranting further investigation.
Tumor cells differ in their tumorigenic potential, metastatic capability, and resistance to treatments. These characteristics are driven by both genetic diversity and cellular plasticity. Increasing evidence indicates that cellular plasticity is closely associated with tumor progression, metastasis, therapy resistance, and relapse and highlights the critical role of ncRNAs in regulating tumor cell plasticity.
Despite the promising advantages of non-invasive early ncRNA detection in liquid biopsies, analyzing various ncRNAs presents specific challenges. For instance, differing miRNA expression levels in blood/serum and solid tumors in the same patient can arise due to different secretion mechanisms or origins [279,280]. The spatio-temporal context of miRNA expression, as reported in previous studies, highlights the difficulty in determining disease site specificity using bulk detection methods.
lncRNAs potentially offer greater specificity due to their generally lower abundance in blood, but face challenges related to their size and stability ratio. These larger molecules are more prone to degradation, making sample integrity a time-sensitive issue, especially in clinical settings.
circRNAs, which are distinguishable from their linear isoforms by the unique sequence created by back-splicing, also present detection challenges. The exclusive junction site can be difficult to detect, and the assumption that the back-splice junction is the sole identifier of circularity can be limiting. Current fragmentation-based next-generation sequencing technologies are not optimal for circRNA detection, and third-generation long-read sequencing methods, while promising, are still in the early stages of establishing themselves as the future gold standard.
Lastly, tRNAs are well-characterized at the sequencing level, but their well-conserved genes can lead to a redundant amino acid coding system. This redundancy may lead to off-target labeling due to the inherent similarities between endogenous tRNA molecules or trigger widespread detrimental effects on protein-coding pathways. The highly conserved nature of tRNA may be its greatest strength or a major limitation in its application for therapeutic purposes, which remains to be investigated.
Given the previously mentioned limitations, the use of ncRNA expression patterns in liquid biopsies for diagnostic, prognostic, and therapeutic purposes is currently being extensively studied in large cancer patient cohorts and more detection methods are being discovered. This research aims to confirm their effectiveness as biomarkers in human cancers.
Analyzing the functions of cancer-related ncRNAs poses challenges, particularly in modulating their expression. Unlike protein-coding genes, where genetic knock-outs can be achieved by introducing a stop codon, lncRNA transcripts themselves are the functional units, complicating knock-out approaches [281–283]. Disrupting core promoters of ncRNA genes is also challenging due to their frequent association with protein-coding genes. Additionally, the efficiency of silencing lncRNAs with siRNAs is often lower than that of proteins because lncRNAs can form stable secondary structures that hinder siRNA binding. Overcoming these challenges requires the development of new strategies to modulate ncRNA expression and the creation of new models for functional studies [284]. These models, like organoids and patient-derived xenografts, can better recapitulate the TME and provide a more accurate assessment of ncRNA functions and their interactions with other cellular components. Future research should focus on identifying novel ncRNAs involved in cellular plasticity and elucidating their mechanisms of action. Emerging technologies, such as single-cell RNA sequencing and CRISPR-based ncRNA editing, will be instrumental in uncovering these regulatory networks. Furthermore, understanding the interplay between different types of ncRNAs and their collective impact on cellular states will provide deeper insights into stem cell biology and cancer. This includes studying the interactions between lncRNAs, miRNAs, and circRNAs, and how they coordinate to regulate gene expression and cellular behavior. Integrative approaches combining transcriptomics, proteomics, and metabolomics will be crucial in mapping the complex regulatory networks governed by ncRNAs.
Another challenge is ensuring the specificity of ncRNA targets involved in key pathways contributing to disease progression, particularly in cancer cell plasticity. The delivery of ncRNA molecules to specific subcellular locations and maintaining their stability are crucial to avoid both “on-target” and “off-target” side effects. The specificity of delivery and treatment towards tumor cells remains a hurdle to overcome. These limitations primarily include insufficient penetration into tumor tissues due to mechanical and biological barriers, ensuring the stability and integrity of ncRNAs in circulation, potential induction of ncRNA-related immunotoxicity, and the risk of off-target effects of ncRNAs.
One effective strategy to mitigate the harmful side effects caused by delivery vehicles is to conjugate the targeting RNA with a ligand that binds to receptors overexpressed in the target cells. This approach enhances cellular uptake without significantly altering biodistribution [285]. Such specificity is particularly advantageous in targeting cancer cells, where ligands like N-acetylgalactosamine can be conjugated to ASOs to bind asialoglycoprotein receptors highly expressed in the liver [286,287].
The development of nanoparticles as delivery vehicles has been a major focus to protect ncRNAs from degradation and enhance their delivery efficiency. However, the formation of a protein corona around nanoparticles in vivo can lead to their preferential uptake by mononuclear phagocytes, which poses a challenge to their effective delivery [288]. Modifying the surface characteristics of NPs, such as PEGylation, can reduce these unwanted interactions and improve targeting efficiency [289]. Rodlike supramolecular nanoassemblies made from degradable poly(aspartic acid) derivatives and hydroxyl-rich cellulose nanocrystals for efficient delivery of tumor-suppressive ncRNAs have also been explored. These nanoassemblies exhibit high gene transfection efficiency and low cytotoxicity, making them a promising platform for cancer therapy due to their biocompatibility and degradability [290]. Furthermore, physiological barriers such as the endothelial and blood-brain barriers, renal clearance, and the reticuloendothelial system need to be addressed to enhance ncRNA delivery. Strategies like the use of ionizable lipids that alter charge properties at different pH levels within endosomes have shown promise in facilitating endosomal escape and improving intracellular delivery [291].
Future research must focus on optimizing these delivery systems and understanding the intricate regulatory networks of ncRNAs in cancer cell plasticity. Advances in nanotechnology, chemical modifications, and bioengineering of delivery vehicles will be fundamental in overcoming these barriers. The integration of artificial intelligence and machine learning could also play a significant role in predicting the interactions of ncRNAs and selecting the most effective therapeutic targets [292].
Non-coding RNAs are pivotal regulators of stem cell and CSC self-renewal and differentiation. Their ability to modulate gene expression and cellular states makes them attractive targets for therapeutic intervention. Continued research into ncRNA functions and mechanisms will enhance our insights into of cellular plasticity and contribute to the development of novel cancer treatments.
Not applicable.
Not applicable.
This work was supported by the Princess Margaret Cancer Foundation (886012001223 to H.H.H.), CIHR operating grants (142246, 152863, 152864 and 159567 to H.H.H.), Terry Fox New Frontiers Program Project Grant (PPG19-1090 and PPG23-1124 to H.H.H.). H.H.H. holds Tier 1 Canada Research Chair in RNA Medicine. P.H. was supported by a MOHCCN Health Informatics Award and a CIHR Doctoral Award.
The authors have declared that no competing interests exist.
| 1. | International Human Genome Sequencing Consortium. Initial sequencing and analysis of the human genome. Nature. 2001;409:860-921. [Google Scholar] [CrossRef] |
| 2. | The ENCODE Project Consortium. Identification and analysis of functional elements in 1% of the human genome by the ENCODE pilot project. Nature. 2007;447:799-816. [Google Scholar] [CrossRef] |
| 3. | Alexander RP, Fang G, Rozowsky J, Snyder M, Gerstein MB. Annotating non-coding regions of the genome. Nat Rev Genet. 2010;11(8):559-571. [Google Scholar] [CrossRef] |
| 4. | Memczak S, Jens M, Elefsinioti A, Torti F, Krueger J, Rybak A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013;495:333-338. [Google Scholar] [CrossRef] |
| 5. | Hansen TB, Jensen TI, Clausen BH, Bramsen JB, Finsen B, Damgaard CK, et al. Natural RNA circles function as efficient microRNA sponges. Nature. 2013;495:384-388. [Google Scholar] [CrossRef] |
| 6. | Liu C-X, Li X, Nan F, Jiang S, Gao X, Guo S-K, et al. Structure and Degradation of Circular RNAs Regulate PKR Activation in Innate Immunity. Cell. 2019;177(4):865-880. [Google Scholar] [CrossRef] |
| 7. | Lyer MK, Niknafs YS, Malik R, Singhal U, Sahu A, Hosono Y, et al. The landscape of long noncoding RNAs in the human transcriptome. Nat Genet. 2015;47(3):199-208. [Google Scholar] [CrossRef] |
| 8. | Thomas RC, Joan AS. The Noncoding RNA Revolution—Trashing Old Rules to Forge New Ones. Cell. 2014;157(1):77-94. [Google Scholar] [CrossRef] |
| 9. | Zhang P, Wu W, Chen Q, Chen M. Non-Coding RNAs and their Integrated Networks. J Integr Bioinform. 2019;16(3):20190027. [Google Scholar] [CrossRef] |
| 10. | Zheng Q, Bao C, Guo W, Li S, Chen J, Chen B, et al. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat Commmun. 2016;7:11215. [Google Scholar] [CrossRef] |
| 11. | Zhong S, Zhou S, Yang S, Yu X, Xu H, Wang J, et al. Identification of internal control genes for circular RNAs. Biotechnol Lett. 2019;41(10):1111-1119. [Google Scholar] [CrossRef] |
| 12. | Kapur M, Ganguly A, Nagy G, Chuang JH, Frankel WN, Ackerman SL. Expression of the Neuronal tRNA n-Tr20 Regulates Synpatic Transmission and Seizure Susceptibility. Neuron. 2020;108(1):193-208. [Google Scholar] [CrossRef] |
| 13. | Cui C, Liu Y, Gerloff D, Rohde C, Pauli C, Köhn M, et al. NOP10 predicts lung cancer prognosis and its associated small nucleolar RNAs drive proliferation and migration. Oncogene. 2020;40(5):909-921. [Google Scholar] [CrossRef] |
| 14. | Monshaugen I, Luna L, Rhodes J, Kristiansen FIS, Lång A, Bøe SO, et al. Depletion of the m1A writer TRMT6/TRMT61A reduces proliferation and resistance against cellular stress in bladder cancer. Front Oncol. 2024;13:1334112. [Google Scholar] [CrossRef] |
| 15. | Cui L, Nakano K, Obchoei S, Setoguchi K, Matsumoto M, Yamamoto T, et al. Small Nucleolar Noncoding RNA SNORA23, Up-Regulated in Human Pancreatic Ductal Adenocarcinoma, Regulates Expression of Spectrin Repeat-Containing Nuclear Envelope 2 to Promote Growth and Metastasis of Xenograft Tumors in Mice. Gastroenterology. 2017;153(1):292-306. [Google Scholar] [CrossRef] |
| 16. | Croce CM. Causes and consequences of microRNA dysregulation in cancer. Nat Rev Genet. 2012;10(10):704-714. [Google Scholar] [CrossRef] |
| 17. | Croce CM, Peng Y. The role of MicroRNAs in human cancer. Signal Transduct Target Ther. 2016;1:15004. [Google Scholar] [CrossRef] |
| 18. | Otmani K, Lewalle P. Tumor Suppressor miRNA in Cancer Cells and the Tumor Microenvironment: Mechanism of Deregulation and Clinical Implications. Front Oncol. 2021;11(1):708765. [Google Scholar] [CrossRef] |
| 19. | Bolatkan A, Asada K, Kaneko S, Suvarna K, Ikawa N, Machino H, et al. Downregulation of METTL6 mitigates cell progression, migration, invasion and adhesion in hepatocellular carcinoma by inhibiting cell adhesion molecules. Int J Oncol. 2021;60:4. [Google Scholar] [CrossRef] |
| 20. | Chiu H-S, Somvanshi S, Patel E, Chen T-W, Singh VP, Zorman B, et al. Pan-Cancer Analysis of lncRNA Regulation Supports Their Targeting of Cancer Genes in Each Tumor Context. Cell Rep. 2018;23(1):297-312.e12. [Google Scholar] [CrossRef] |
| 21. | Taniue K, Akimitsu N. The Functions and Unique Features of LncRNAs in Cancer Development and Tumorigenesis. Int J Mol Sci. 2021;22(2):632. [Google Scholar] [CrossRef] |
| 22. | Jiang M-C, Ni J-J, Cui W-Y, Wang B-Y, Zhuo W. Emerging roles of lncRNA in cancer and therapeutic opportunities. Am J Cancer Res. 2019;9(7):1354-1366. [Google Scholar] |
| 23. | Vo JN, Cieslik M, Zhang Y, Shukla S, Xiao L, Zhang Y, et al. The Landscape of Circular RNA in Cancer. Cell. 2019;176(4):869-881. [Google Scholar] [CrossRef] |
| 24. | Pisignano G, Michael DC, Visal TH, Pirlog R, Ladomery M, Calin GA. Going circular: history, present, and future of circRNAs in cancer. Oncogene. 2023;42(38):2783-2800. [Google Scholar] [CrossRef] |
| 25. | Xia H, Li X, Gao W, Fu X, Fang RH, Zhang L, et al. Tissue repair and regeneration with endogenous stem cells. Nat Rev Mater. 2018;3(1):174-193. [Google Scholar] [CrossRef] |
| 26. | Lu Y, Brommer B, Tian X, Krishnan A, Meer M, Wang C, et al. Reprogramming to recover youthful epigenetic information and restore vision. Nature. 2020;588(7836):124-129. [Google Scholar] [CrossRef] |
| 27. | Lu L, Finegold MJ, Johnson RL. Hippo pathway coactivators Yap and Taz are required to coordinate mammlian liver regeneration. Exp Mol Med. 2018;50(1):e423. [Google Scholar] [CrossRef] |
| 28. | Shah M, Allegrucci C. Stem Cell Plasticity in Development and Cancer: Epigenetic Origin of Cancer Stem Cells. Subcell Biochem. 2012;61(1):545-565. [Google Scholar] [CrossRef] |
| 29. | Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF. Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci U S A. 2003;100(7):3983-3988. [Google Scholar] [CrossRef] |
| 30. | Singh SK, Clarke ID, Terasaki M, Bonn VE, Hawkins C, Squire J, et al. Identification of a Cancer Stem Cell in Human Brain Tumors. Cancer Res. 2003;63(18):5821-5828. [Google Scholar] |
| 31. | Fang D, Nguyen TK, Leishear K, Finko R, Kulp AN, Hotz S, et al. A Tumorigenic Subpopulation with Stem Cell Properties in Melanomas. Cancer Res. 2005;65(20):9328-9337. [Google Scholar] [CrossRef] |
| 32. | Hermann PC, Huber SL, Herrler T, Aicher A, Ellwart JW, Guba M, et al. Distinct Populations of Cancer Stem Cells Determine Tumor Growth and Metastatic Activity in Human Pancreatic Cancer. Cell Stem Cell. 2007;1(3):313-323. [Google Scholar] [CrossRef] |
| 33. | Gu G, Yuan J, Wills M, Kasper S. Prostate cancer cells with stem cell characteristics reconstitute the original human tumor in vivo. Cancer Res. 2007;67(10):4807-4815. [Google Scholar] [CrossRef] |
| 34. | Bapat SA, Mali AM, Koppikar CB, Kurrey NK. Stem and Progenitor-Like Cells Contribute to the Aggressive Behavior of Human Epithelial Ovarian Cancer. Cancer Res. 2005;65(8):3025-3029. [Google Scholar] [CrossRef] |
| 35. | Luginbühl J, Sivaraman DM, Shin JW. The essentiality of non-coding RNAs in cell reprogramming. Noncoding RNA Res. 2017;2(1):74-82. [Google Scholar] [CrossRef] |
| 36. | Xu J, Liu S. Noncoding RNAs in Cancer Cell Plasticity. Adv Exp Med Biol. 2016;927:173-189. [Google Scholar] [CrossRef] |
| 37. | Ghildiyal M, Zamore PD. Small silencing RNAs: an expanding universe. Nat Rev Genet. 2009;10(2):94-108. [Google Scholar] [CrossRef] |
| 38. | O'Brien J, Hayder H, Zayed Y, Peng C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front Endocrinol (Lausanne). 2018;9(1):402. [Google Scholar] [CrossRef] |
| 39. | Kirstein N, Dokaneheifard S, Cingaram PR, Valencia MG, Beckedorff F, Santos HGD, et al. The Integrator complex regulates microRNA abundance through RISC loading. Sci Adv. 2023;9(6):eadf0597. [Google Scholar] [CrossRef] |
| 40. | Kozomara A, Birgaoanu M, Griffiths-Jones S. miRBase: from microRNA sequences to function. Nucleic Acids Res. 2019;47(D1):D155-D62. [Google Scholar] [CrossRef] |
| 41. | Kent OA, Chivukula RR, Mullendore M, Wentzel EA, Feldmann G, Lee KH, et al. Repression of the miR-143/145 cluster by oncogenic Ras initiates a tumor-promoting feed-forward pathway. Genes Dev. 2010;24(24):2754-2759. [Google Scholar] [CrossRef] |
| 42. | Roderburg C, Luedde T. Circulating microRNAs as markers of liver inflammation, fibrosis and cancer. J Hepatol. 2014;61(6):1434-1437. [Google Scholar] [CrossRef] |
| 43. | Yanaihara N, Caplen N, Bowman E, Seike M, Kumamoto K, Yi M, et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell. 2006;9(3):189-198. [Google Scholar] [CrossRef] |
| 44. | Calin GA, Ferracin M, Cimmino A, Di Leva G, Shimizu M, Wojcik SE, et al. A MicroRNA Signature Associated with Prognosis and Progression in Chronic Lymphocytic Leukemia. N Engl J Med. 2005;353(17):1793-1801. [Google Scholar] [CrossRef] |
| 45. | Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J, Peck D, et al. MicroRNA expression profiles classify human cancers. Nature. 2005;435(7043):834-838. [Google Scholar] [CrossRef] |
| 46. | Carter JV, Galbraith NJ, Yang D, Burton JF, Walker SP, Galandiuk S. Blood-based microRNAs as biomarkers for the diagnosis of colorectal cancer: a systematic review and meta-analysis. Br J Cancer. 2017;116(6):762-774. [Google Scholar] [CrossRef] |
| 47. | Mitchell PS, Parkin RK, Kroh EM, Fritz BR, Wyman SK, Pogosova-Agadjanyan EL, et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl Acad Sci USA. 2008;105(30):10513-10518. [Google Scholar] [CrossRef] |
| 48. | Kopp F, Mendell JT. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell. 2018;172(3):393-407. [Google Scholar] [CrossRef] |
| 49. | Rinn JL, Chang HY. Genome Regulation by Long Noncoding RNAs. Annu Rev Biochem. 2013;81(1):145-166. [Google Scholar] [CrossRef] |
| 50. | Derrien T, Johnson R, Bussotti G, Tanzer A, Djebali S, Tilgner H, et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012;22(9):1775-1789. [Google Scholar] [CrossRef] |
| 51. | Sartorelli V, Lauberth SM. Enhancer RNAs are an important regulatory layer of the epigenome. Nat Struct Mol Biol. 2020;27(6):521-528. [Google Scholar] [CrossRef] |
| 52. | Zhang M, Zhao J, Wu J, Wang Y, Zhuang M, Zou L, et al. In-depth characterization and identification of translatable lncRNAs. Comput Biol Med. 2023;164:107243. [Google Scholar] [CrossRef] |
| 53. | Uszczynska-Ratajczak B, Lagarde J, Frankish A, Guigó R, Johnson R. Towards a complete map of the human long non-coding RNA transcriptome. Nat Rev Gen. 2018;19(9):535-548. [Google Scholar] [CrossRef] |
| 54. | Fang S, Zhang L, Guo J, Niu Y, Wu Y, Li H, et al. NONCODEV5: a comprehensive annotation database for long non-coding RNAs. Nucleic Acids Res. 2018;46(D1):D308-D314. [Google Scholar] [CrossRef] |
| 55. | Sánchez Y, Segura V, Marín-Béjar O, Athie A, Marchese FP, González J, et al. Genome-wide analysis of the human p53 transcriptional network unveils a lncRNA tumour suppressor signature. Nat Commun. 2014;5:5812. [Google Scholar] [CrossRef] |
| 56. | Huarte M, Guttman M, Feldser D, Garber M, Koziol MJ, Kenzelmann-Broz D, et al. A Large Intergenic Noncoding RNA Induced by p53 Mediates Global Gene Repression in the p53 Response. Cell. 2010;142(3):409-419. [Google Scholar] [CrossRef] |
| 57. | Hart JR, Roberts TC, Weinberg MS, Morris KV, Vogt PK. MYC regulates the non-coding transcriptome. Oncotarget. 2014;5(24):12543-12554. [Google Scholar] [CrossRef] |
| 58. | Kim T, Jeon Y-J, Cui R, Lee J-H, Peng Y, Kim S-H, et al. Role of MYC-Regulated Long Noncoding RNAs in Cell Cycle Regulation and Tumorigenesis. J Natl Cancer Inst. 2015;107(4):dju505. [Google Scholar] [CrossRef] |
| 59. | Chakravarty D, Sboner A, Nair SS, Giannopoulou E, Li R, Hennig S, et al. The oestrogen receptor alpha-regulated lncRNA NEAT1 is a critical modulator of prostate cancer. Nature Commun. 2014;5:5383. [Google Scholar] [CrossRef] |
| 60. | Trimarchi T, Bilal E, Ntziachristos P, Fabbri G, Dalla-Favera R, Tsirigos A, et al. Genome-wide Mapping and Characterization of Notch-Regulated Long Noncoding RNAs in Acute Leukemia. Cell. 2014;158(3):593-606. [Google Scholar] [CrossRef] |
| 61. | Sharma U, Barwal TS, Malhotra A, Pant N, Vivek Dey D, et al. Long non-coding RNA TINCR as potential biomarker and therapeutic target for cancer. Life Sci. 2020 Sep 15;257:118035. [Google Scholar] [CrossRef] |
| 62. | Zhang Y, Zhang X-O, Chen T, Xiang J-F, Yin Q-F, Xing Y-H, et al. Circular Intronic Long Noncoding RNAs. Mol Cell. 2013;51(6):792-806. [Google Scholar] [CrossRef] |
| 63. | Zhang X-O, Wang H-B, Zhang Y, Lu X, Chen L-L, Yang L. Complementary Sequence-Mediated Exon Circularization. Cell. 2014;159(1):134-147. [Google Scholar] [CrossRef] |
| 64. | Ivanov A, Memczak S, Wyler E, Torti F, Hagit Marta, et al. Analysis of Intron Sequences Reveals Hallmarks of Circular RNA Biogenesis in Animals. Cell Rep. 2015;10(2):170-177. [Google Scholar] [CrossRef] |
| 65. | Ashwal-Fluss R, Meyer M, Nagarjuna Ivanov A, Bartok O, Hanan M, et al. circRNA Biogenesis Competes with Pre-mRNA Splicing. Mol Cell. 2014;56(1):55-66. [Google Scholar] [CrossRef] |
| 66. | Conn SJ, Pillman KA, Toubia J, Conn VM, Salmanidis M, Phillips CA, et al. The RNA Binding Protein Quaking Regulates Formation of circRNAs. Cell. 2015;160(6):1125-1134. [Google Scholar] [CrossRef] |
| 67. | Li X, Liu C-X, Xue W, Zhang Y, Jiang S, Yin Q-F, et al. Coordinated circRNA Biogenesis and Function with NF90/NF110 in Viral Infection. Mol Cell. 2017;67(2):214-227. [Google Scholar] [CrossRef] |
| 68. | Liang D, Tatomer DC, Luo Z, Wu H, Yang L, Chen L-L, et al. The Output of Protein-Coding Genes Shifts to Circular RNAs When the Pre-mRNA Processing Machinery Is Limiting. Mol Cell. 2017;68(5):940-954. [Google Scholar] [CrossRef] |
| 69. | Kramer MC, Liang D, Tatomer DC, Gold B, March ZM, Cherry S, et al. Combinatorial control of Drosophila circular RNA expression by intronic repeats, hnRNPs, and SR proteins. Genes Dev. 2015;29(20):2168-2182. [Google Scholar] [CrossRef] |
| 70. | Liang D, Wilusz JE. Short intronic repeat sequences facilitate circular RNA production. Genes Dev. 2014;28(20):2233-2247. [Google Scholar] [CrossRef] |
| 71. | Glažar P, Papavasileiou P, Rajewsky N. circBase: a database for circular RNAs. RNA. 2014;20(11):1666-1670. [Google Scholar] [CrossRef] |
| 72. | Ji P, Wu W, Chen S, Zheng Y, Zhou L, Zhang J, et al. Expanded Expression Landscape and Prioritization of Circular RNAs in Mammals. Cell Rep. 2019;26(12):3444-3460. [Google Scholar] [CrossRef] |
| 73. | Wu W, Ji P, Zhao F. CircAtlas: an integrated resource of one million highly accurate circular RNAs from 1070 vertebrate transcriptomes. Genome Biol. 2020;21:101. [Google Scholar] [CrossRef] |
| 74. | Dong R, Ma X-K, Li G-W, Yang L. CIRCpedia v2: An Updated Database for Comprehensive Circular RNA Annotation and Expression Comparison. Genomics Proteomoics Bioinformatics. 2018;16(4):226-233. [Google Scholar] [CrossRef] |
| 75. | Dahl M, Husby S, Eskelund CW, Besenbacher S, Fjelstrup S, Côme C, et al. Expression patterns and prognostic potential of circular RNAs in mantle cell lymphoma: a study of younger patients from the MCL2 and MCL3 clinical trials. Leukemia. 2022;36:177-188. [Google Scholar] [CrossRef] |
| 76. | Geng Y, Jiang J, Wu C. Function and clinical significance of circRNAs in solid tumors. J Hematol Oncol. 2018;11:98. [Google Scholar] [CrossRef] |
| 77. | Huijbers A, Tollenaar RAEM, V Pelt GW, Zeestraten ECM, Dutton S, McConkey CC, et al. The proportion of tumor-stroma as a strong prognosticator for stage II and III colon cancer patients: validation in the VICTOR trial. Ann Oncol. 2012;24:179-185. [Google Scholar] [CrossRef] |
| 78. | Gujam FJA, Edwards J, Mohammed ZMA, Going JJ, McMillan DC. The relationship between the tumour stroma percentage, clinicopathological characteristics and outcome in patients with operable ductal breast cancer. Br J Cancer. 2014;111:157-165. [Google Scholar] [CrossRef] |
| 79. | Ahmadov U, Bendikas MM, Ebbesen KK, Sehested AM, Kjems J, Broholm H, et al. Distinct circular RNA expression profiles in pediatric ependymomas. Brain Pathol. 2021;31(2):387-392. [Google Scholar] [CrossRef] |
| 80. | Rickert D, Bartl J, Picard D, Bernardi F, Qin N, Lovino M, et al. Circular RNA profiling distinguishes medulloblastoma groups and shows aberrant RMST overexpression in WNT medulloblastoma. Acta Neuropatholo. 2021;141(6):975-978. [Google Scholar] [CrossRef] |
| 81. | Nair AA, Niu N, Tang X, Thompson KJ, Wang L, Kocher J-P, et al. Circular RNAs and their associations with breast cancer subtypes. Oncotarget. 2016;7(49):80967-80979. [Google Scholar] [CrossRef] |
| 82. | Wang C, Tan S, Liu W-R, Lei Q, Qiao W, Wu Y, et al. RNA-Seq profiling of circular RNA in human lung adenocarcinoma and squamous cell carcinoma. Mol Cancer. 2019;18:134. [Google Scholar] [CrossRef] |
| 83. | Suzuki T, Yashiro Y, Kikuchi I, Ishigami Y, Saito H, Matsuzawa I, et al. Complete chemical structures of human mitochondrial tRNAs. Nat Commun. 2020;11:4269. [Google Scholar] [CrossRef] |
| 84. | Chatterjee K, Nostramo RT, Wan Y, Hopper AK. tRNA dynamics between the nucleus, cytoplasm and mitochondrial surface: Location, location, location. Biochim Biophys Acta. 2017;1861(4):373-386. [Google Scholar] [CrossRef] |
| 85. | Parisien M, Wang X, Pan T. Diversity of human tRNA genes from the 1000-genomes project. RNA Biol. 2013;10(12):1853-1867. [Google Scholar] [CrossRef] |
| 86. | Agris PF, Vendeix FAP, Graham WD. tRNA’s Wobble Decoding of the Genome: 40 Years of Modification. J Mol Biol. 2007;366(1):1-13. [Google Scholar] [CrossRef] |
| 87. | Berg MD, Brandl CJ. Transfer RNAs: diversity in form and function. RNA Biol. 2020;18(3):316-339. [Google Scholar] [CrossRef] |
| 88. | Clark WC, Evans ME, Dominissini D, Zheng G, Pan T. tRNA base methylation identification and quantification via high-throughput sequencing. RNA. 2016;22(11):1771-1784. [Google Scholar] [CrossRef] |
| 89. | Pan T. Modifications and functional genomics of human transfer RNA. Cell Res. 2018;28:395-404. [Google Scholar] [CrossRef] |
| 90. | Ivanov P, Emara MM, Villen J, Gyqi SP, Anderson P. Angiogenin-induced tRNA Fragments Inhibit Translation Initiation. Mol Cell. 2011;43(4):613-623. [Google Scholar] [CrossRef] |
| 91. | Honda S, Loher P, Shigematsu M, Palazzo JP, Suzuki R, Imoto I, et al. Sex hormone-dependent tRNA halves enhance cell proliferation in breast and prostate cancers. Proc Natl Acad Sci USA. 2015;112(29):E3816-E3825. [Google Scholar] [CrossRef] |
| 92. | Chen Z, Qi M, Shen B, Luo G, Wu Y, Li J, et al. Transfer RNA demethylase ALKBH3 promotes cancer progression via induction of tRNA-derived small RNAs. Nucleic Acids Res. 2019;47(5):2533-2545. [Google Scholar] [CrossRef] |
| 93. | Zakrzewski W, Dobrzyński M, Szymonowicz M, Rybak Z. Stem cells: past, present, and future. Stem Cell Res Ther. 2019;10(1):68. [Google Scholar] [CrossRef] |
| 94. | Subramanyam D, Lamouille S, Judson RL, Liu JY, Bucay N, Derynck R, et al. Multiple targets of miR-302 and miR-372 promote reprogramming of human fibroblasts to induced pluripotent stem cells. Nat Biotech. 2011;29(5):443-448. [Google Scholar] [CrossRef] |
| 95. | Li Z, Yang C-S, Nakashima K, Rana TM. Small RNA-mediated regulation of iPS cell generation. EMBO J. 2011;30(5):823-834. [Google Scholar] [CrossRef] |
| 96. | Pfaff N, Liebhaber S, Möbus S, Beh-Pajooh A, Fiedler J, Pfanne A, et al. Inhibition of miRNA-212/132 improves the reprogramming of fibroblasts into induced pluripotent stem cells by de-repressing important epigenetic remodelling factors. Stem Cell Res. 2017;20(1):70-75. [Google Scholar] [CrossRef] |
| 97. | Vencken SF, Sethupathy P, Blackshields G, Spillane C, Elbaruni S, Sheils O, et al. An integrated analysis of the SOX2 microRNA response program in human pluripotent and nullipotent stem cell lines. BMC Genomics. 2014;15(1):711. [Google Scholar] [CrossRef] |
| 98. | Xu N, Papagiannakopoulos T, Pan G, Thomson JA, Kosik KS. MicroRNA-145 regulates OCT4, SOX2, and KLF4 and represses pluripotency in human embryonic stem cells. Cell. 2009;137(4):647-658. [Google Scholar] [CrossRef] |
| 99. | Rosa A, Brivanlou AH. A regulatory circuitry comprised of miR-302 and the transcription factors OCT4 and NR2F2 regulates human embryonic stem cell differentiation. EMBO J. 2011;30(2):237-248. [Google Scholar] [CrossRef] |
| 100. | Hui M, Lu Y, Zhang Z, Chen S, Qi Y, Xu Y, et al. MicroRNA-21 maintains hematopoietic stem cell homeostasis through sustaining the NF-κB signaling pathway in mice. Haematologica. 2021;106(2):412-423. [Google Scholar] [CrossRef] |
| 101. | Kurkewich JL, Boucher A, Klopfenstein N, Baskar R, Kapur R, Dahl R. The mirn23a and mirn23b microrna clusters are necessary for proper hematopoietic progenitor cell production and differentiation. Exp Hematol. 2018;59:14-29. [Google Scholar] [CrossRef] |
| 102. | Zhao JL, Rao DS, O'Connell RM, Garcia-Flores Y, Baltimore D. MicroRNA-146a acts as a guardian of the quality and longevity of hematopoietic stem cells in mice. Elife. 2013;2:e00537. [Google Scholar] [CrossRef] |
| 103. | Schulman RMS, Esquela-Kerscher A, Slack FJ. Reciprocal expression of lin-41 and the microRNAs let-7 and mir-125 during mouse embryogenesis. Dev Dyn. 2005;234(4):1046-1054. [Google Scholar] [CrossRef] |
| 104. | Copley MR, Babovic S, Benz C, Knapp DJH, Beer AB, Kent DG. The Lin28b-let-7-Hmga2 axis determines the higher self-renewal potential of fetal haematopoietic stem cells. Nat Cell Biol. 2013;15(8):916-925. [Google Scholar] [CrossRef] |
| 105. | Lee YT, Vasconcellos JFd, Yuan J, Byrnes C, Noh S-J, et al. LIN28B-mediated expression of fetal hemoglobin and production of fetal-like erythrocytes from adult human erythroblasts ex vivo. Blood. 2013;122(6):1034-1041. [Google Scholar] [CrossRef] |
| 106. | Emmrich S, Rasche M, Schöning J, Reimer C, Keihani S, Maroz A, et al. miR-99a/100~125b tricistrons regulate hematopoietic stem and progenitor cell homeostasis by shifting the balance between TGFβ and Wnt signaling. Genes Dev. 2014;28(8):858-874. [Google Scholar] [CrossRef] |
| 107. | Shi Y, Zhao X, Hsieh J, Wichterle H, Impey S, Banerjee S, et al. MicroRNA Regulation of Neural Stem Cells and Neurogenesis. J Neurosci. J Neurosci. 2010;30(45):14931-14936. [Google Scholar] [CrossRef] |
| 108. | Zhao C, Sun G, Li S, Shi Y. A feedback regulatory loop involving microRNA-9 and nuclear receptor TLX in neural stem cell fate determination. Nat Struct Mol Biol. 2009;16(4):365-371. [Google Scholar] [CrossRef] |
| 109. | Mirzadeh Azad F, Polignano IL, Proserpio V, Oliviero S. Long Noncoding RNAs in Human Stemness and Differentiation. Trends in Cell Biology. 2021;31(7):542-555. [Google Scholar] [CrossRef] |
| 110. | Chakraborty D, Paszkowski-Rogacz M, Berger N, Ding L, Mircetic J, Fu J, et al. lncRNA Panct1 Maintains Mouse Embryonic Stem Cell Identity by Regulating TOBF1 Recruitment to Oct-Sox Sequences in Early G1. Cell Rep. 2017;21(11):3012-3021. [Google Scholar] [CrossRef] |
| 111. | Brown CJ, Ballabio A, Rupert JL, Lafreniere RG, Grompe M, Tonlorenzi R, et al. A gene from the region of the human X inactivation centre is expressed exclusively from the inactive X chromosome. Nature. 1991;349(6304):38-44. [Google Scholar] [CrossRef] |
| 112. | Chelmicki T, Dündar F, Turley MJ, Khanam T, Aktas T, Ramírez F, et al. MOF-associated complexes ensure stem cell identity and Xist repression. eLife. 2014;3:e02024. [Google Scholar] [CrossRef] |
| 113. | An C, Feng G, Zhang J, Lu F, Zhou Q, Wang H, et al. Overcoming Autocrine FGF Signaling-Induced Heterogeneity in Naive Human ESCs Enables Modeling of Random X Chromosome Inactivation. Cell Stem Cell. 2020;27(3):482-497. [Google Scholar] [CrossRef] |
| 114. | Li W, Ren Y, Si Y, Wang F, Yu J. Long non-coding RNAs in hematopoietic regulation. Cell Regen. 2018;7(2):27-32. [Google Scholar] [CrossRef] |
| 115. | Venkatraman A, He XC, Thorvaldsen JL, Sugimura R, Perry JM, Tao F, et al. Maternal imprinting at the H19-Igf2 locus maintains adult haematopoietic stem cell quiescence. Nature. 2013;500(7462):345-349. [Google Scholar] [CrossRef] |
| 116. | Bond AM, Vangompel MJW, Sametsky EA, Clark MF, Savage JC, Disterhoft JF, et al. Balanced gene regulation by an embryonic brain ncRNA is critical for adult hippocampal GABA circuitry. Nat Neurosci. 2009;12(8):1020-1027. [Google Scholar] [CrossRef] |
| 117. | Ramos AD, Andersen RE, Liu SJ, Nowakowski TJ, Hong SJ, Gertz C, et al. The long noncoding RNA Pnky regulates neuronal differentiation of embryonic and postnatal neural stem cells. Cell Stem Cell. 2015;16(4):439-447. [Google Scholar] [CrossRef] |
| 118. | Ninou E, Michail A, Politis PK. Long Non-Coding RNA Lacuna Regulates Neuronal Differentiation of Neural Stem Cells During Brain Development. Front Cell Devl Biol. 2021;9(1):726857. [Google Scholar] [CrossRef] |
| 119. | Lu H-J, Li J, Yang G, Yi C-J, Zhang D, Yu F, et al. Circular RNAs in stem cells: from basic research to clinical implications. Biosci Rep. 2022;42(1):BSR20212510. [Google Scholar] [CrossRef] |
| 120. | Yu C-Y, Li T-C, Wu Y-Y, Yeh C-H, Chiang W, Chuang C-Y, et al. The circular RNA circBIRC6 participates in the molecular circuitry controlling human pluripotency. Nat Commun. 2017;8:1149. [Google Scholar] [CrossRef] |
| 121. | Errichelli L, Modigliani SD, Laneve P, Colantoni A, Legnini I, Capauto D, et al. FUS affects circular RNA expression in murine embryonic stem cell-derived motor neurons. Nat Commun. 2017;30(8):14741. [Google Scholar] [CrossRef] |
| 122. | Xia P, Wang S, Ye B, Du Y, Li C, Xiong Z, et al. A Circular RNA Protects Dormant Hematopoietic Stem Cells from DNA Sensor cGAS-Mediated Exhaustion. Immunity. 2018;48(4):688-701.e7. [Google Scholar] [CrossRef] |
| 123. | Nicolet BP, Jansen SBG, Heideveld E, Ouwehand WH, Akker EMVD, Lindern MV, et al. Circular RNAs exhibit limited evidence for translation, or translation regulation of the mRNA counterpart in terminal hematopoiesis. RNA. 2022;28(2):194-209. [Google Scholar] [CrossRef] |
| 124. | Nicolet BP, Engels S, Aglialoro F, Akker EMVD, Lindern MV, Wolkers MC. Circular RNA expression in human hematopoietic cells is widespread and cell-type specific. Nucleic Acids Res. 2018;46(16):8168-8180. [Google Scholar] [CrossRef] |
| 125. | Yang Q, Wu J, Zhao J, Xu T, Zhao Z, Song X, Han P. Circular RNA expression profiles during the differentiation of mouse neural stem cells. BMC Syst Biol. 2018;12:128. [Google Scholar] [CrossRef] |
| 126. | Seeler S, Andersen MS, Sztanka-Toth T, Rybiczka-Tešulov M, Munkhof MHvd, Chang C-C, et al. A Circular RNA Expressed from the FAT3 Locus Regulates Neural Development. Mol Neurobiol. 2023;6(6):3239-3260. [Google Scholar] [CrossRef] |
| 127. | Wang G, Han B, Shen L, Wu S, Yang L, Liao J, et al. Silencing of circular RNA HIPK2 in neural stem cells enhances functional recovery following ischaemic stroke. EBioMedicine. 2020;52:102660. [Google Scholar] [CrossRef] |
| 128. | Saba JA, Liakath-Ali K, Green R, Watt FM. Translational control of stem cell function. Nat Rev Mol Cell Biol. 2021;22(10):671-690. [Google Scholar] [CrossRef] |
| 129. | Krishna S, Yim DG, Lakshmanan V, Tirumalai V, Koh JL, Park JE, et al. Dynamic expression of tRNA-derived small RNAs define cellular states. EMBO Rep. 2019;20(7):e47789. [Google Scholar] [CrossRef] |
| 130. | Sobala A, Hutvagner G. Small RNAs derived from the 5′ end of tRNA can inhibit protein translation in human cells. RNA Biol. 2013;10(4):553-563. [Google Scholar] [CrossRef] |
| 131. | Weidensdorfer D, Stöhr N, Baude A, Lederer M, Köhn M, Schierhorn A, et al. Control of c-myc mRNA stability by IGF2BP1-associated cytoplasmic RNPs. RNA. 2009;15(1):104-115. [Google Scholar] [CrossRef] |
| 132. | Lambrianidou A, Sereti E, Soupsana K, Komini C, Dimas K, Trangas T. mTORC2 deploys the mRNA binding protein IGF2BP1 to regulate c-MYC expression and promote cell survival. Cellular Signal. 2021;80:109912. [Google Scholar] [CrossRef] |
| 133. | Schwartz S, Douglas AB, Maxwell RM, Jovanovic M, Herbst RH, León-Ricardo B, et al. Transcriptome-wide Mapping Reveals Widespread Dynamic-Regulated Pseudouridylation of ncRNA and mRNA. Cell. 2014;159:148-162. [Google Scholar] [CrossRef] |
| 134. | Guzzi N, Cieśla M, Ngoc PCT, Lang S, Arora S, Dimitriou M, et al. Pseudouridylation of tRNA-Derived Fragments Steers Translational Control in Stem Cells. Cell. 2018;173(5):1204-1216.e26. [Google Scholar] [CrossRef] |
| 135. | Zhang F, Yoon K, Zhang DY, Kim N-S, Ming G-L, Song H. Epitranscriptomic regulation of cortical neurogenesis via Mettl8-dependent mitochondrial tRNA m3C modification. Cell Stem Cell. 2023;30(3):300-311.e11. [Google Scholar] [CrossRef] |
| 136. | Carey LA, Perou CM, Livasy CA, Dressler LG, Cowan D, Conway K, et al. Race, Breast Cancer Subtypes, and Survival in the Carolina Breast Cancer Study. JAMA. 2006;295(21):2492-2502. [Google Scholar] [CrossRef] |
| 137. | Dravis C, Chung C-Y, Lytle NK, Herrera-Valdez J, Luna G, Trejo CL, et al. Epigenetic and Transcriptomic Profiling of Mammary Gland Development and Tumor Models Disclose Regulators of Cell State Plasticity. Cancer Cell. 2018;34(3):466-482.e6. [Google Scholar] [CrossRef] |
| 138. | Mohamed GA, Mahmood S, Ognjenovic NB, Lee MK, Wilkins OM, Christensen BC, et al. Lineage plasticity enables low-ER luminal tumors to evolve and gain basal-like traits. Breast Cancer Res. 2023;25(1):23. [Google Scholar] [CrossRef] |
| 139. | Al-Masri M, Paliotti K, Tran R, Halaoui R, Lelarge V, Chatterjee S, et al. Architectural control of metabolic plasticity in epithelial cancer cells. Commun Biol. 2021;4:371. [Google Scholar] [CrossRef] |
| 140. | Mukha A, Kahya U, Linge A, Chen O, Löck S, Lukiyanchuk V, et al. GLS-driven glutamine catabolism contributes to prostate cancer radiosensitivity by regulating the redox state, stemness and ATG5-mediated autophagy. Theranostics. 2021;11(16):7844-7868. [Google Scholar] [CrossRef] |
| 141. | Bhat GR, Sethi I, Sadida HQ, Rah B, Mir R, Algehainy N, et al. Cancer cell plasticity: from cellular, molecular, and genetic mechanisms to tumor heterogeneity and drug resistance. Cancer Metastasis Rev. 2024;43(1):197-228. [Google Scholar] [CrossRef] |
| 142. | Khan A, Ahmed E, Elareer N, Junejo K, Steinhoff M, Uddin S. Role of miRNA-Regulated Cancer Stem Cells in the Pathogenesis of Human Malignancies. Cells. 2019;8(8):840. [Google Scholar] [CrossRef] |
| 143. | Yan H, Bu P. Non-coding RNAs in cancer stem cells. Cancer Lett. 2018;421:121-126. [Google Scholar] [CrossRef] |
| 144. | Al-Sowayan BS, Al-Shareeda AT, Alrfaei BM. Cancer Stem Cell-Exosomes, Unexposed Player in Tumorigenicity. Front Pharmacol. 2020;11(1):384. [Google Scholar] [CrossRef] |
| 145. | Guessous F, Zhang Y, Kofman A, Catania A, Li Y, Schiff D, et al. microRNA-34a is tumor suppressive in brain tumors and glioma stem cells. Cell Cycle. 2010;9(6):1031-1036. [Google Scholar] [CrossRef] |
| 146. | Liu C, Kelnar K, Liu B, Chen X, Calhoun-Davis T, Li H, et al. The microRNA miR-34a inhibits prostate cancer stem cells and metastasis by directly repressing CD44. Nat Med. 2011;17(2):211-215. [Google Scholar] [CrossRef] |
| 147. | Shimono Y, Zabala M, Cho RW, Lobo N, Dalerba P, Qian D, et al. Downregulation of miRNA-200c Links Breast Cancer Stem Cells with Normal Stem Cells. Cell. 2009;138(3):592-603. [Google Scholar] [CrossRef] |
| 148. | Iliopoulos D, Lindahl-Allen M, Polytarchou C, Hirsch HA, Tsichlis PN, Struhl K. Loss of miR-200 Inhibition of Suz12 Leads to Polycomb-Mediated Repression Required for the Formation and Maintenance of Cancer Stem Cells. Mol Cell. 2010;39(5):761-772. [Google Scholar] [CrossRef] |
| 149. | Lim YY, Wright JA, Attema JL, Gregory PA, Bert AG, Smith E, et al. Epigenetic modulation of the miR-200 family is associated with transition to a breast cancer stem cell-like state. J Cell Sci. 2013;126(10):2256-2266. [Google Scholar] [CrossRef] |
| 150. | Zhang L, Li H, Zang Y, Wang F. NLRP3 inflammasome inactivation driven by miR-223-3p reduces tumor growth and increases anticancer immunity in breast cancer. Mol Med Rep. 2019;19(3):2180-2188. [Google Scholar] [CrossRef] |
| 151. | Liu C, Liu R, Zhang D, Deng Q, Liu B, Chao H-P, et al. MicroRNA-141 suppresses prostate cancer stem cells and metastasis by targeting a cohort of pro-metastasis genes. Nat Commun. 2017;8:14270. [Google Scholar] [CrossRef] |
| 152. | Xu D-D, Zhou P-J, Wang Y, Zhang Y, Zhang R, Zhang L, et al. miR-150 Suppresses the Proliferation and Tumorigenicity of Leukemia Stem Cells by Targeting the Nanog Signaling Pathway. Front Pharmacol. 2016;7:439. [Google Scholar] [CrossRef] |
| 153. | Yan T-T, Ren L-L, Shen C-Q, Wang Z-H, Yu Y-N, Liang Q, et al. miR-508 Defines the Stem-like/Mesenchymal Subtype in Colorectal Cancer. Cancer Res. 2018;78(7):1751-1765. [Google Scholar] [CrossRef] |
| 154. | Cai H, Chen Y, Yang X, Ma S, Wang Q, Zhang Y, et al. Let7b modulates the Wnt/β-catenin pathway in liver cancer cells via downregulated Frizzled4. Tumor Biol. 2017;39(7):101042831771607. [Google Scholar] [CrossRef] |
| 155. | Jin B, Wang W, Meng X-X, Du G, Li J, Zhang S-Z, et al. Let-7 inhibits self-renewal of hepatocellular cancer stem-like cells through regulating the epithelial-mesenchymal transition and the Wnt signaling pathway. BMC Cancer. 2016;16:863. [Google Scholar] [CrossRef] |
| 156. | Yu F, Yao H, Zhu P, Zhang X, Pan Q, Gong C, et al. let-7 Regulates Self Renewal and Tumorigenicity of Breast Cancer Cells. Cell. 2007;131(6):1109-1123. [Google Scholar] [CrossRef] |
| 157. | Lv C, Li F, Li X, Tian Y, Zhang Y, Sheng X, et al. MiR-31 promotes mammary stem cell expansion and breast tumorigenesis by suppressing Wnt signaling antagonists. Nat Commun. 2017;8:1036. [Google Scholar] [CrossRef] |
| 158. | Ren J, Kuang T-H, Chen J, Yang J-W, Liu Y-X. The diagnostic and prognostic values of microRNA-21 in patients with gastric cancer: a meta-analysis. Eur Rev Med Pharmacol Sci. 2017;21(1):120-130. [Google Scholar] |
| 159. | Ouzounova M, Vuong T, Ancey P-B, Ferrand M, Durand G, Le-Calvez Kelm F, et al. MicroRNA miR-30 family regulates non-attachment growth of breast cancer cells. BMC Genomics. 2013;14(1):139. [Google Scholar] [CrossRef] |
| 160. | Zhang S, He Y, Liu C, Li G, Lu S, Jing Q, et al. miR-93-5p enhances migration and invasion by targeting RGMB in squamous cell carcinoma of the head and neck. J Cancer. 2020;11(13):3871-3881. [Google Scholar] [CrossRef] |
| 161. | Li N, Miao Y, Shan Y, Liu B, Li Y, Zhao L, et al. MiR-106b and miR-93 regulate cell progression by suppression of PTEN via PI3K/Akt pathway in breast cancer. Cell Death Dis. 2017;8(5):e2796. [Google Scholar] [CrossRef] |
| 162. | Hudson RS, Yi M, Esposito D, Glyn SA, Starks AM, Yang Y, et al. MicroRNA-106b-25 cluster expression is associated with early disease recurrence and targets caspase-7 and focal adhesion in human prostate cancer. Oncogene. 2013;32(35):4139-4147. [Google Scholar] [CrossRef] |
| 163. | Xu C, Shi L, Chen W, Fang P, Li J, Jin L, et al. MiR-106b inhibitors sensitize TRAIL-induced apoptosis in hepatocellular carcinoma through increase of death receptor 4. Oncotarget. 2017;8(26):41921-41931. [Google Scholar] [CrossRef] |
| 164. | Zhang G-J, Li L-F, Yang G-D, Xia S-S, Wang R, Leng Z-W, et al. MiR-92a promotes stem cell-like properties by activating Wnt/β-catenin signaling in colorectal cancer. Oncotarget. 2017;8(60):101760-101770. [Google Scholar] [CrossRef] |
| 165. | Jiang C, Yu M, Xie X, Huang G, Peng Y, Ren D, et al. miR-217 targeting DKK1 promotes cancer stem cell properties via activation of the Wnt signaling pathway in hepatocellular carcinoma. Oncol Rep. 2017;38(4):2351-2359. [Google Scholar] [CrossRef] |
| 166. | Zheng Z, Liu J, Yang Z, Wu L, Xie H, Jiang C, et al. MicroRNA-452 promotes stem-like cells of hepatocellular carcinoma by inhibiting Sox7 involving Wnt/β-catenin signaling pathway. Oncotarget. 2016;7(19):28000-28012. [Google Scholar] [CrossRef] |
| 167. | Du J, Liu S, He J, Liu X, Qu Y, Yan W, et al. MicroRNA-451 regulates stemness of side population cells via PI3K/Akt/mTOR signaling pathway in multiple myeloma. Oncotarget. 2015;6(17):14993-15007. [Google Scholar] [CrossRef] |
| 168. | Su JS, Ito K, Ala U, Kats L, Webster K, Sun SM, et al. The Oncogenic MicroRNA miR-22 Targets the TET2 Tumor Suppressor to Promote Hematopoietic Stem Cell Self-Renewal and Transformation. Cell Stem Cell. 2013;13(1):87-101. [Google Scholar] [CrossRef] |
| 169. | Title AC, Hong S-J, Pires ND, Hasenöhrl L, Godbersen S, Stokar-Regenscheit N, et al. Genetic dissection of the miR-200–Zeb1 axis reveals its importance in tumor differentiation and invasion. Nat Commun. 2018;9:4671. [Google Scholar] [CrossRef] |
| 170. | Diaz-Riascos ZV, Ginesta MM, Fabregat J, Buscail L, Cordelier P, Capellá G. Expression and Role of MicroRNAs from the miR-200 Family in the Tumor Formation and Metastatic Propensity of Pancreatic Cancer. Mol Ther Nucleic Acids. 2019;17(1):491-503. [Google Scholar] [CrossRef] |
| 171. | Gregory PA, Bert AG, Paterson EL, Barry SC, Tsykin A, Farshid G, et al. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat Cell Biol. 2008;10(5):593-601. [Google Scholar] [CrossRef] |
| 172. | Chen S, Zhu J, Wang F, Guan Z, Ge Y, Yang X, et al. LncRNAs and their role in cancer stem cells. Oncotarget. 2017;8(66):110685-110692. [Google Scholar] [CrossRef] |
| 173. | Duan Y, Jia Y, Wang J, Liu T, Cheng Z, Sang M, et al. Long noncoding RNA DGCR5 involves in tumorigenesis of esophageal squamous cell carcinoma via SRSF1-mediated alternative splicing of Mcl-1. Cell Death Dis. 2021;12(6):587. [Google Scholar] [CrossRef] |
| 174. | Ji Q, Zhang L, Liu X, Zhou L, Wang W, Han Z, et al. Long non-coding RNA MALAT1 promotes tumour growth and metastasis in colorectal cancer through binding to SFPQ and releasing oncogene PTBP2 from SFPQ/PTBP2 complex. Br J Cancer. 2014;111(4):736-748. [Google Scholar] [CrossRef] |
| 175. | Meng L-D, Shi G-D, Ge W-L, Huang X-M, Cheng Q, Yuan H, et al. Linc01232 promotes the metastasis of pancreatic cancer by suppressing the ubiquitin-mediated degradation of HNRNPA2B1 and activating the A-Raf-induced MAPK/ERK signaling pathway. Cancer Lett. 2020;494:107-120. [Google Scholar] [CrossRef] |
| 176. | Chen Y-T, Kan C-H, Liu H, Liu Y-H, Wu C-C, Kuo Y-P, et al. Modular scaffolding by lncRNA HOXA10-AS promotes oral cancer progression. Cell Death Dis. 2022;13(7):629. [Google Scholar] [CrossRef] |
| 177. | Shao M, Yang Q, Zhu W, Jin H, Wang J, Song J, et al. LncHOXA10 drives liver TICs self-renewal and tumorigenesis via HOXA10 transcription activation. Mol Cancer. 2018;17:173. [Google Scholar] [CrossRef] |
| 178. | Wang Y, Zhu P, Luo J, Wang J, Liu Z, Wu W, et al. LncRNA HAND2-AS1 promotes liver cancer stem cell self-renewal via BMP signaling. EMBO J. 2019;38(17):e101110. [Google Scholar] [CrossRef] |
| 179. | Wang X, Sun W, Shen W, Xia M, Chen C, Xiang D, et al. Long non-coding RNA DILC regulates liver cancer stem cells via IL-6/STAT3 axis. J Hepatol. 2016;64(6):1283-1294. [Google Scholar] [CrossRef] |
| 180. | Wang J, Lei Z-J, Guo Y, Wang T, Qin Z-Y, Xiao H-L, et al. miRNA-regulated delivery of lincRNA-p21 suppresses β-catenin signaling and tumorigenicity of colorectal cancer stem cells. Oncotarget. 2015;6(35):37852-37870. [Google Scholar] [CrossRef] |
| 181. | Chen Q, Liu X, Xu L, Wang Y, Wang S, Li Q, et al. Long non-coding RNA BACE1-AS is a novel target for anisomycin-mediated suppression of ovarian cancer stem cell proliferation and invasion. Oncol Rep. 2016;35(4):1916-1924. [Google Scholar] [CrossRef] |
| 182. | Chen X, Xie R, Gu P, Huang M, Han J, Dong W, et al. Long Noncoding RNA LBCS Inhibits Self-Renewal and Chemoresistance of Bladder Cancer Stem Cells through Epigenetic Silencing of SOX2. Clin Cancer Res. 2019;25(4):1389-1403. [Google Scholar] [CrossRef] |
| 183. | Ma Y, Zhu Y, Shang L, Qiu Y, Shen N, Wang J, et al. LncRNA XIST regulates breast cancer stem cells by activating proinflammatory IL-6/STAT3 signaling. Oncogene. 2023;42():1419-1437. [Google Scholar] [CrossRef] |
| 184. | Bauderlique-Le Roy H, Vennin C, Brocqueville G, Spruyt N, Adriaenssens E, Bourette RP. Enrichment of Human Stem-Like Prostate Cells with s-SHIP Promoter Activity Uncovers a Role in Stemness for the Long Noncoding RNA H19. Stem Cells Dev. 2015;24(10):1252-1262. [Google Scholar] [CrossRef] |
| 185. | Peng F, Li T-T, Wang K-L, Xiao G-Q, Wang J-H, Zhao H-D, et al. H19/let-7/LIN28 reciprocal negative regulatory circuit promotes breast cancer stem cell maintenance. Cell Death Dis. 2017;8:e2569. [Google Scholar] [CrossRef] |
| 186. | Jiao F, Hu H, Han T, Yuan C, Wang L, Jin Z, et al. Long Noncoding RNA MALAT-1 Enhances Stem Cell-Like Phenotypes in Pancreatic Cancer Cells. Int J Mol Sci. 2015;16(4):6677-6693. [Google Scholar] [CrossRef] |
| 187. | Alves PC, Fonseca AS, Muys BR, de Barros E Lima Bueno R, Bürger MC, Souza JES, et al. Brief Report: The lincRNA Hotair Is Required for Epithelial-to-Mesenchymal Transition and Stemness Maintenance of Cancer Cell Lines. Stem Cells. 2013;31(12):2827-2832. [Google Scholar] [CrossRef] |
| 188. | Zhang H, Cai K, Wang J, Wang X, Cheng K, Shi F, et al. MiR-7, Inhibited Indirectly by LincRNA HOTAIR, Directly Inhibits SETDB1 and Reverses the EMT of Breast Cancer Stem Cells by Downregulating the STAT3 Pathway. Stem Cells. 2014;32(11):2858-2868. [Google Scholar] [CrossRef] |
| 189. | Ma M-Z, Li C-X, Zhang Y, Weng M-Z, Zhang M-d, Qin Y-Y, et al. Long non-coding RNA HOTAIR, a c-Myc activated driver of malignancy, negatively regulates miRNA-130a in gallbladder cancer. Mol Cancer. 2014;13(1):156. [Google Scholar] [CrossRef] |
| 190. | Li Z, Jiang P, Li J, Peng M, Zhao X, Zhang X, et al. Tumor-derived exosomal lnc-Sox2ot promotes EMT and stemness by acting as a ceRNA in pancreatic ductal adenocarcinoma. Oncogene. 2018;37(28):3822-3838. [Google Scholar] [CrossRef] |
| 191. | Zhang J-J, Zhu Y, Zhang X-F, Liu D-F, Wang Y, Yang C, et al. Yin Yang-1 suppresses pancreatic ductal adenocarcinoma cell proliferation and tumor growth by regulating SOX2OT-SOX2 axis. Cancer Lett. 2017;408(1):144-154. [Google Scholar] [CrossRef] |
| 192. | Gui X, Li H, Li T, Pu H, Lu D. Long Noncoding RNA CUDR Regulates HULC and β-Catenin to Govern Human Liver Stem Cell Malignant Differentiation. Mol Ther. 2015;23(12):1843-1853. [Google Scholar] [CrossRef] |
| 193. | Wu J, Zhu P, Lu T, Du Y, Wang Y, He L, et al. The long non-coding RNA LncHDAC2 drives the self-renewal of liver cancer stem cells via activation of Hedgehog signaling. J Hepatol. 2018;70(5):918-929. [Google Scholar] [CrossRef] |
| 194. | Wang Y, He L, Du Y, Zhu P, Huang G, Luo J, et al. The Long Noncoding RNA lncTCF7 Promotes Self-Renewal of Human Liver Cancer Stem Cells through Activation of Wnt Signaling. Cell Stem Cell. 2015;16(4):413-425. [Google Scholar] [CrossRef] |
| 195. | Qu L, Wu Z, Li Y, Xu Z, Liu B, Liu F, et al. A feed-forward loop between lncARSR and YAP activity promotes expansion of renal tumour-initiating cells. Nat Commun. 2016;7:12692. [Google Scholar] [CrossRef] |
| 196. | Chen Z-Z, Huang L, Wu Y-H, Zhai W-J, Zhu P-P, Gao Y-F. LncSox4 promotes the self-renewal of liver tumour-initiating cells through Stat3-mediated Sox4 expression. Nat Commun. 2016;7:12598. [Google Scholar] [CrossRef] |
| 197. | Yuan SX, Wang J, Yang F, Tao QF, Zhang J, Wang LL, et al. Long noncoding RNA DANCR increases stemness features of hepatocellular carcinoma by derepression of CTNNB1. Hepatology. 2016;63(2):499-511. [Google Scholar] [CrossRef] |
| 198. | Zhu P, Wang Y, Huang G, Ye B, Liu B, Wu J, et al. lnc-β-Catm elicits EZH2-dependent β-catenin stabilization and sustains liver CSC self-renewal. Nat Struct Mol Biol. 2016;23(7):631-639. [Google Scholar] [CrossRef] |
| 199. | Wu M, Lin Z, Li X, Xin X, An J, Zheng Q, et al. HULC cooperates with MALAT1 to aggravate liver cancer stem cells growth through telomere repeat-binding factor 2. Sci Rep. 2016;6:36045. [Google Scholar] [CrossRef] |
| 200. | Tripathi V, Shen Z, Chakraborty A, Giri S, Freier SM, Wu X, et al. Long Noncoding RNA MALAT1 Controls Cell Cycle Progression by Regulating the Expression of Oncogenic Transcription Factor B-MYB. PLoS Genet. 2013;9(3):e1003368. [Google Scholar] [CrossRef] |
| 201. | Askarian-Amiri ME, Seyfoddin V, Smart CE, Wang J, Kim JE, Hansji H, et al. Emerging Role of Long Non-Coding RNA SOX2OT in SOX2 Regulation in Breast Cancer. PLoS ONE. 2014;9(7):e102140. [Google Scholar] [CrossRef] |
| 202. | Ashrafizadeh M, Dai J, Torabian P, Nabavi N, Aref AR, Aljabali AAA, et al. Circular RNAs in EMT-driven metastasis regulation: modulation of cancer cell plasticity, tumorigenesis and therapy resistance. CMLS. 2024;81(1):214. [Google Scholar] [CrossRef] |
| 203. | Zhu Y-J, Zheng B, Luo G-J, Ma X-K, Lu X-Y, Lin X-M, et al. Circular RNAs negatively regulate cancer stem cells by physically binding FMRP against CCAR1 complex in hepatocellular carcinoma. Theranostics. 2019;9(12):3526-3540. [Google Scholar] [CrossRef] |
| 204. | Lin X, Chen W, Wei F, Xie X. TV-circRGPD6 Nanoparticle Suppresses Breast Cancer Stem Cell-Mediated Metastasis via the miR-26b/YAF2 Axis. Mol Ther. 2021;29(1):244-262. [Google Scholar] [CrossRef] |
| 205. | Zhang L, Dong X, Yan B, Yu W, Shan L. CircAGFG1 drives metastasis and stemness in colorectal cancer by modulating YY1/CTNNB1. Cell Death Dis. 2020;11(7):542. [Google Scholar] [CrossRef] |
| 206. | Li W, Yang X, Shi C, Zhou Z. Hsa_circ_002178 Promotes the Growth and Migration of Breast Cancer Cells and Maintains Cancer Stem-like Cell Properties Through Regulating miR-1258/KDM7A Axis. Cell Transplant. 2020;29:096368972096017. [Google Scholar] [CrossRef] |
| 207. | Zhan W, Liao X, Wang Y, Li L, Li J, Chen Z, et al. circCTIC1 promotes the self-renewal of colon TICs through BPTF-dependent c-Myc expression. Carcinogenesis. 2018;40(4):560-568. [Google Scholar] [CrossRef] |
| 208. | Gao X, Xia X, Li F, Zhang M, Zhou A, Wu X, et al. Circular RNA-encoded oncogenic E-cadherin variant promotes glioblastoma tumorigenicity through activation of EGFR-STAT3 signalling. Nat Cell Biol. 2021;28:278-291. [Google Scholar] [CrossRef] |
| 209. | Rengganaten V, Huang C-J, Wang M-L, Chien Y, Tsai P-H, Lan Y-T, et al. Circular RNA ZNF800 (hsa_circ_0082096) regulates cancer stem cell properties and tumor growth in colorectal cancer. BMC Cancer. 2023;23:1088. [Google Scholar] [CrossRef] |
| 210. | Li C, Zhang J, Yang X, Hu C, Chu T, Zhong R, et al. hsa_circ_0003222 accelerates stemness and progression of non-small cell lung cancer by sponging miR-527. Cell Death Dis. 2021;12:807. [Google Scholar] [CrossRef] |
| 211. | Jian X, He H, Zhu J, Zhang Q, Zheng Z, Liang X, et al. Correction: Hsa_circ_001680 affects the proliferation and migration of CRC and mediates its chemoresistance by regulating BMI1 through miR-340. Mol Cancer. 2024;23:100. [Google Scholar] [CrossRef] |
| 212. | Li X, Liu X, Zhao D, Cui W, Wu Y, Zhang C, et al. tRNA-derived small RNAs: novel regulators of cancer hallmarks and targets of clinical application. Cell Death Dis. 2021;7:249. [Google Scholar] [CrossRef] |
| 213. | Feng W, Li Y, Chu J, Li J, Zhang Y, Ding X, et al. Identification of tRNA-derived small noncoding RNAs as potential biomarkers for prediction of recurrence in triple-negative breast cancer. Cancer Med. 2018;7(10):5130-5144. [Google Scholar] [CrossRef] |
| 214. | Mo D, He F, Zheng J, Chen H, Tang L, Yan F. tRNA-Derived Fragment tRF-17-79MP9PP Attenuates Cell Invasion and Migration via THBS1/TGF-β1/Smad3 Axis in Breast Cancer. Front Oncol. 2021;11:656078. [Google Scholar] [CrossRef] |
| 215. | Huang B, Yang H, Cheng X, Wang D, Fu S, Shen W, et al. tRF/miR-1280 Suppresses Stem Cell–like Cells and Metastasis in Colorectal Cancer. Cancer Res. 2017;77(12):3194-3206. [Google Scholar] [CrossRef] |
| 216. | Zhou Y, Hu J, Liu L, Yan M, Zhang Q, Song X, et al. Gly-tRF enhances LCSC-like properties and promotes HCC cells migration by targeting NDFIP2. Cancer Cell Int. 2021;21:502. [Google Scholar] [CrossRef] |
| 217. | Marrugo-Ramírez J, Mir M, Samitier J. Blood-Based Cancer Biomarkers in Liquid Biopsy: A Promising Non-Invasive Alternative to Tissue Biopsy. Int J Mol Sci. 2018;19(10):2877. [Google Scholar] [CrossRef] |
| 218. | Sestini S, Boeri M, Marchiano A, Pelosi G, Galeone C, Verri C, et al. Circulating microRNA signature as liquid-biopsy to monitor lung cancer in low-dose computed tomography screening. Oncotarget. 2015;6(32):32868-32877. [Google Scholar] [CrossRef] |
| 219. | De Fraipont F, Gazzeri S, Cho WC, Eymin B. Circular RNAs and RNA Splice Variants as Biomarkers for Prognosis and Therapeutic Response in the Liquid Biopsies of Lung Cancer Patients. Front Genet. 2019;10:390. [Google Scholar] [CrossRef] |
| 220. | Lampis A, Ghidini M, Ratti M, Mirchev MB, Okuducu AF, Valeri N, et al. Circulating Tumour DNAs and Non-Coding RNAs as Liquid Biopsies for the Management of Colorectal Cancer Patients. Gastrointest Disord. 2020;2(3):212-235. [Google Scholar] [CrossRef] |
| 221. | Badowski C, He B, Garmire LX. Blood-derived lncRNAs as biomarkers for cancer diagnosis: the Good, the Bad and the Beauty. NPJ Precis Oncol. 2022;6:40. [Google Scholar] [CrossRef] |
| 222. | Wang J, Ma G, Ge H, Han X, Mao X, Wang X, et al. Circulating tRNA-derived small RNAs (tsRNAs) signature for the diagnosis and prognosis of breast cancer. NPJ Breast Cancer. 2021;7:4. [Google Scholar] [CrossRef] |
| 223. | Shigeyasu K, Toden S, Zumwalt TJ, Okugawa Y, Goel A. Emerging Role of MicroRNAs as Liquid Biopsy Biomarkers in Gastrointestinal Cancers. Clin Cancer Res. 2017;23(10):2391-2399. [Google Scholar] [CrossRef] |
| 224. | Leong SM, Tan M-L, Chua HW, Huang M-C, Cheong WC, Li M-H, et al. Paper-Based MicroRNA Expression Profiling from Plasma and Circulating Tumor Cells. Clin Chem. 2017;63(3):731-741. [Google Scholar] [CrossRef] |
| 225. | Markou A, Zavridou M, Sourvinou I, Yousef G, Kounelis S, Malamos N, et al. Direct Comparison of Metastasis-Related miRNAs Expression Levels in Circulating Tumor Cells, Corresponding Plasma, and Primary Tumors of Breast Cancer Patients. Clin Chem. 2016;62(7):1002-1011. [Google Scholar] [CrossRef] |
| 226. | Shoshan E, Mobley AK, Braeuer RR, Kamiya T, Huang L, Vasquez ME, et al. Reduced adenosine-to-inosine miR-455-5p editing promotes melanoma growth and metastasis. Nat Cell Biol. 2015;17(3):311-321. [Google Scholar] [CrossRef] |
| 227. | Melo SA, Luecke LB, Kahlert C, Fernandez AF, Gammon ST, Kaye J, et al. Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature. 2015;523(7559):177-182. [Google Scholar] [CrossRef] |
| 228. | Rane JK, Scaravilli M, Ylipää A, Pellacani D, Mann VM, Simms MS, et al. MicroRNA Expression Profile of Primary Prostate Cancer Stem Cells as a Source of Biomarkers and Therapeutic Targets. Eur Urol. 2015;67(1):7-10. [Google Scholar] [CrossRef] |
| 229. | Taylor BS, Schultz N, Hieronymus H, Gopalan A, Xiao Y, Carver BS, et al. Integrative Genomic Profiling of Human Prostate Cancer. Cancer Cell. 2010;18(1):11-22. [Google Scholar] [CrossRef] |
| 230. | Collins AT, Berry PA, Hyde C, Stower MJ, Maitland NJ. Prospective Identification of Tumorigenic Prostate Cancer Stem Cells. Cancer Res. 2005;65(23):10946-10951. [Google Scholar] [CrossRef] |
| 231. | Richardson GD, Robson CN, Lang SH, Neal DE, Maitland NJ, Collins AT. CD133, a novel marker for human prostatic epithelial stem cells. J Cell Sci. 2004;117(16):3539-3545. [Google Scholar] [CrossRef] |
| 232. | Chi H-C, Tsai C-Y, Tsai M-M, Yeh C-T, Lin K-H. Roles of Long Noncoding RNAs in Recurrence and Metastasis of Radiotherapy-Resistant Cancer Stem Cells. Int J Mol Sci. 2017;18(9):1903. [Google Scholar] [CrossRef] |
| 233. | Terracciano D, Ferro M, Terreri S, Lucarelli G, D'Elia C, Musi G, et al. Urinary long noncoding RNAs in nonmuscle-invasive bladder cancer: new architects in cancer prognostic biomarkers. Transl Res. 2017;184:108-117. [Google Scholar] [CrossRef] |
| 234. | Lemos AEG, Matos AdR, Ferreira LB, Gimba ERP. The long non-coding RNA PCA3: an update of its functions and clinical applications as a biomarker in prostate cancer. Oncotarget. 2019;10(61):6589-6603. [Google Scholar] [CrossRef] |
| 235. | Song T, Xu A, Zhang Z, Gao F, Zhao L, Chen X, et al. CircRNA hsa_circRNA_101996 increases cervical cancer proliferation and invasion through activating TPX2 expression by restraining miR-8075. J Cell Physiol. 2019;234(8):14296-14305. [Google Scholar] [CrossRef] |
| 236. | Endo H, Shiroki T, Nakagawa T, Yokoyama M, Tamai K, Yamanami H, et al. Enhanced Expression of Long Non-Coding RNA HOTAIR Is Associated with the Development of Gastric Cancer. PLoS ONE. 2013;8(10):e77070. [Google Scholar] [CrossRef] |
| 237. | Lisowski L, Dane AP, Chu K, Zhang Y, Cunningham SC, Wilson EM, et al. Selection and evaluation of clinically relevant AAV variants in a xenograft liver model. Nature. 2014;506(7488):382-386. [Google Scholar] [CrossRef] |
| 238. | Hang D, Zhou J, Qin N, Zhou W, Ma H, Jin G, et al. A novel plasma circular RNA circFARSA is a potential biomarker for non-small cell lung cancer. Cancer Med. 2018;7(6):2783-2791. [Google Scholar] [CrossRef] |
| 239. | Li Y, Zheng Q, Bao C, Li S, Guo W, Zhao J, et al. Circular RNA is enriched and stable in exosomes: a promising biomarker for cancer diagnosis. Cell Res. 2015;25(8):981-984. [Google Scholar] [CrossRef] |
| 240. | Wen N, Peng D, Xiong X, Liu G, Nie G, Wang Y, et al. Cholangiocarcinoma combined with biliary obstruction: an exosomal circRNA signature for diagnosis and early recurrence monitoring. Signal Transduct Target Ther. 2024;9(1):107. [Google Scholar] [CrossRef] |
| 241. | Ruggero D. Translational Control in Cancer Etiology. Cold Spring Harb Perspect Biol. 2013;5(2):a012336. [Google Scholar] [CrossRef] |
| 242. | Liu W, Yu M, Cheng S, Zhou X, Li J, Lu Y, et al. tRNA-Derived RNA Fragments Are Novel Biomarkers for Diagnosis, Prognosis, and Tumor Subtypes in Prostate Cancer. Curr Oncol. 2023;30(1):981-999. [Google Scholar] [CrossRef] |
| 243. | Kasinski AL, Slack FJ. Epigenetics and genetics. MicroRNAs en route to the clinic: progress in validating and targeting microRNAs for cancer therapy. Nat Rev Cancer. 2011;11(12):849-864. [Google Scholar] [CrossRef] |
| 244. | Lennox KA, Behlke MA. A direct comparison of anti-microRNA oligonucleotide potency. Pharm Res. 2010 Apr 28;27(1):1788-1799. [Google Scholar] [CrossRef] |
| 245. | Si M-L, Zhu S, Wu H, Lu Z, Wu F, Mo Y-Y. miR-21-mediated tumor growth. Oncogene. 2007;26(19):2799-2803. [Google Scholar] [CrossRef] |
| 246. | Wang J, Li Y, Ding M, Zhang H, Xu X, Tang J. Molecular mechanisms and clinical applications of miR-22 in regulating malignant progression in human cancer (Review). Int J Oncol. 2016;50(2):345-355. [Google Scholar] [CrossRef] |
| 247. | Kyriazi AA, Papiris E, Kalyvianakis KK, Sakellaris G, Baritaki S. Dual Effects of Non-Coding RNAs (ncRNAs) in Cancer Stem Cell Biology. Int J Mol Sci. 2020;21(18):6658. [Google Scholar] [CrossRef] |
| 248. | Zuber J, McJunkin K, Fellmann C, Dow LE, Taylor MJ, Hannon GJ, et al. Toolkit for evaluating genes required for proliferation and survival using tetracycline-regulated RNAi. Nat Biotechnol. 2011;29(1):79-83. [Google Scholar] [CrossRef] |
| 249. | Watanabe C, Cuellar TL, Haley B. Quantitative evaluation of first, second, and third generation hairpin systems reveals the limit of mammalian vector-based RNAi. RNA Biol. 2016;13(1):25-33. [Google Scholar] [CrossRef] |
| 250. | Arun G, Diermeier S, Akerman M, Chang K-C, Wilkinson JE, Hearn S, et al. Differentiation of mammary tumors and reduction in metastasis upon Malat1 lncRNA loss. Genes Dev. 2016;30(1):34-51. [Google Scholar] [CrossRef] |
| 251. | Gutschner T, Hämmerle M, Eißmann M, Hsu J, Kim Y, Hung G, et al. The Noncoding RNA MALAT1 Is a Critical Regulator of the Metastasis Phenotype of Lung Cancer Cells. Cancer Res. 2013;73(3):1180-1189. [Google Scholar] [CrossRef] |
| 252. | Koch L. Functional genomics: Screening for lncRNA function. Nat Rev Genet. 2017;18(2):70. [Google Scholar] [CrossRef] |
| 253. | Gilbert LA, Horlbeck MA, Adamson B, Jacqueline Chen Y, Whitehead EH, et al. Genome-Scale CRISPR-Mediated Control of Gene Repression and Activation. Cell. 2014;159(3):647-661. [Google Scholar] [CrossRef] |
| 254. | Adriaens C, Standaert L, Barra J, Latil M, Verfaillie A, Kalev P, et al. p53 induces formation of NEAT1 lncRNA-containing paraspeckles that modulate replication stress response and chemosensitivity. Nat Med. 2016;22(8):861-868. [Google Scholar] [CrossRef] |
| 255. | Mendell JT. Targeting a Long Noncoding RNA in Breast Cancer. N Engl J Med. 2016;374(23):2287-2289. [Google Scholar] [CrossRef] |
| 256. | Yang H, Wang H, Chikdu Albert, Shi L, Jaenisch R. One-Step Generation of Mice Carrying Reporter and Conditional Alleles by CRISPR/Cas-Mediated Genome Engineering. Cell. 2013;154(6):1370-1379. [Google Scholar] [CrossRef] |
| 257. | Cho SW, Kim S, Kim Y, Kweon J, Kim HS, Bae S, et al. Analysis of off-target effects of CRISPR/Cas-derived RNA-guided endonucleases and nickases. Genome Res. 2014;24(1):132-141. [Google Scholar] [CrossRef] |
| 258. | Bainbridge JWB, Smith AJ, Barker SS, Robbie S, Henderson R, Balaggan K, et al. Effect of Gene Therapy on Visual Function in Leber's Congenital Amaurosis. N Engl J Med. 2008;358(21):2231-2239. [Google Scholar] [CrossRef] |
| 259. | Schaffer DV, Koerber JT, Lim K-I. Molecular Engineering of Viral Gene Delivery Vehicles. Annu Rev Biomed Eng. 2008;10:169-194. [Google Scholar] [CrossRef] |
| 260. | Meganck RM, Borchardt EK, Castellanos Rivera RM, Scalabrino ML, Wilusz JE, Marzluff WF, et al. Tissue-Dependent Expression and Translation of Circular RNAs with Recombinant AAV Vectors In Vivo. Mol Ther Nucleic Acids. 2018;13:89-98. [Google Scholar] [CrossRef] |
| 261. | Han D, Wang Y, Wang Y, Dai X, Zhou T, Chen J, et al. The Tumor-Suppressive Human Circular RNA CircITCH Sponges miR-330-5p to Ameliorate Doxorubicin-Induced Cardiotoxicity Through Upregulating SIRT6, Survivin, and SERCA2a. Circ Res. 2020;127(4):e108-e125. [Google Scholar] [CrossRef] |
| 262. | Rossbach O. Artificial Circular RNA Sponges Targeting MicroRNAs as a Novel Tool in Molecular Biology. Mol Ther Nucleic Acids. 2019;17:452-454. [Google Scholar] [CrossRef] |
| 263. | Kristensen LS, Jakobsen T, Hager H, Kjems J. The emerging roles of circRNAs in cancer and oncology. Nat Rev Clin Oncol. 2022;19(3):188-206. [Google Scholar] [CrossRef] |
| 264. | He AT, Liu J, Li F, Yang BB. Targeting circular RNAs as a therapeutic approach: current strategies and challenges. Signal Transduct Target Ther. 2021;6(1):185. [Google Scholar] [CrossRef] |
| 265. | Wang AZ, Langer R, Farokhzad OC. Nanoparticle Delivery of Cancer Drugs. Annu Rev Med. 2012;63(1):185-198. [Google Scholar] [CrossRef] |
| 266. | Tam Y, Chen S, Cullis P. Advances in Lipid Nanoparticles for siRNA Delivery. Pharmaceutics. 2013;5(3):498-507. [Google Scholar] [CrossRef] |
| 267. | Li D, Gao C, Kuang M, Xu M, Wang B, Luo Y, et al. Nanoparticles as Drug Delivery Systems of RNAi in Cancer Therapy. Molecules. 2021;26(8):2380. [Google Scholar] [CrossRef] |
| 268. | Oliveira ACN, Fernandes J, Gonçalves A, Gomes AC, Oliveira MECDR. Lipid-based Nanocarriers for siRNA Delivery: Challenges, Strategies and the Lessons Learned from the DODAX: MO Liposomal System. Curr Drug Targets. 2019;20(1):29-50. [Google Scholar] [CrossRef] |
| 269. | Fang L, Du WW, Awan FM, Dong J, Yang BB. The circular RNA circ-Ccnb1 dissociates Ccnb1/Cdk1 complex suppressing cell invasion and tumorigenesis. Cancer Lett. 2019;459:216-226. [Google Scholar] [CrossRef] |
| 270. | Rapino F, Delaunay S, Rambow F, Zhou Z, Tharun L, De Tullio P, et al. Codon-specific translation reprogramming promotes resistance to targeted therapy. Nature. 2018;558(7711):605-609. [Google Scholar] [CrossRef] |
| 271. | Chen HM, Wang J, Zhang YF, Gao YH. Ovarian cancer proliferation and apoptosis are regulated by human transfer RNA methyltransferase 9-likevia LIN9. Oncol Lett. 2017;14(4):4461-4466. [Google Scholar] [CrossRef] |
| 272. | Yoshida T, Morihiro K, Naito Y, Mikami A, Kasahara Y, Inuoue T, et al. Identification of nucleobase chemical modifications that reduce the hepatotoxicity of gapmer antisense oligonucleotides. Nucleic Acids Res. 2022;50(13):7224-7234. [Google Scholar] [CrossRef] |
| 273. | Hagedorn PH, Yakimov V, Ottosen S, Kammler S, Nielsen NF, Høg AM, et al. Hepatotoxic potential of therapeutic oligonucleotides can be predicted from their sequence and modification pattern. Nucleic Acid Ther. 2013;23(5):302-310. [Google Scholar] [CrossRef] |
| 274. | Sarli SL, Fakih HH, Kelly K, Devi G, Rembetsy-Brown JM, McEachern HR, et al. Quantifying the activity profile of ASO and siRNA conjugates in glioblastoma xenograft tumors in vivo. Nucleic Acids Res. 2024;52(9):4799-4817. [Google Scholar] [CrossRef] |
| 275. | Hong DS, Kang Y-K, Borad M, Sachdev J, Ejadi S, Lim HY, et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br J Cancer. 2020;122(11):1630-1637. [Google Scholar] [CrossRef] |
| 276. | Grimm D, Streetz KL, Jopling CL, Storm TA, Pandey K, Davis CR, et al. Fatality in mice due to oversaturation of cellular microRNA/short hairpin RNA pathways. Nature. 2006;441(7092):537-541. [Google Scholar] [CrossRef] |
| 277. | Bhan A, Soleimani M, Mandal SS. Long Noncoding RNA and Cancer: A New Paradigm. Cancer Res. 2017;77(15):3965-3981. [Google Scholar] [CrossRef] |
| 278. | Tang Q. Swei. HOTAIR: An Oncogenic Long Non-Coding RNA in Human Cancer. Cell Physiol Biochem. 2018;47(3):893-913. [Google Scholar] [CrossRef] |
| 279. | Lodes MJ, Caraballo M, Suciu D, Munro S, Kumar A, Anderson B. Detection of Cancer with Serum miRNAs on an Oligonucleotide Microarray. PLoS ONE. 2009;4(7):e6229. [Google Scholar] [CrossRef] |
| 280. | Gmerek L, Martyniak K, Horbacka K, Krokowicz P, Scierski W, Golusinski P, et al. MicroRNA regulation in colorectal cancer tissue and serum. PLoS ONE. 2019;14(8):e0222013. [Google Scholar] [CrossRef] |
| 281. | Fathi Dizaji B. Strategies to target long non-coding RNAs in cancer treatment: progress and challenges. EJMHG. 2020;21(1):41. [Google Scholar] [CrossRef] |
| 282. | Grillone K, Riillo C, Scionti F, Rocca R, Tradigo G, Guzzi PH, et al. Non-coding RNAs in cancer: platforms and strategies for investigating the genomic “dark matter”. J Exp Clin Cancer Res. 2020;39(1):117. [Google Scholar] [CrossRef] |
| 283. | Wang J, He X, Corpe C. Molecular Mechanisms and Clinical Implications of Noncoding RNAs in Cancer. Non-Coding RNA. [Google Scholar] [CrossRef] |
| 284. | Sun Y-M, Chen Y-Q. Principles and innovative technologies for decrypting noncoding RNAs: from discovery and functional prediction to clinical application. J Hematol Oncol. 2020;13(1):109. [Google Scholar] [CrossRef] |
| 285. | Bander NH. Antibody–Drug Conjugate Target Selection: Critical Factors. Methods Mol Biol. 2013;1045(1):29-40. [Google Scholar] [CrossRef] |
| 286. | Orellana EA, Tenneti S, Rangasamy L, Lyle LT, Low PS, Kasinski AL. FolamiRs: Ligand-targeted, vehicle-free delivery of microRNAs for the treatment of cancer. Sci Transl Med. 2017;9(401):eaam9327. [Google Scholar] [CrossRef] |
| 287. | Viney NJ, Van Capelleveen JC, Geary RS, Xia S, Tami JA, Yu RZ, et al. Antisense oligonucleotides targeting apolipoprotein(a) in people with raised lipoprotein(a): two randomised, double-blind, placebo-controlled, dose-ranging trials. The Lancet. 2016;388(10057):2239-2253. [Google Scholar] [CrossRef] |
| 288. | Piella J, Bastús NG, Puntes V. Size-Dependent Protein–Nanoparticle Interactions in Citrate-Stabilized Gold Nanoparticles: The Emergence of the Protein Corona. Bioconjug Chem. 2017;28(1):88-97. [Google Scholar] [CrossRef] |
| 289. | Gheibihayat SM, Jaafari MR, Hatamipour M, Sahebkar A. Improvement of the pharmacokinetic characteristics of liposomal doxorubicin using CD47 biomimickry. J Pharm Pharmacol. 2021;73(2):169-177. [Google Scholar] [CrossRef] |
| 290. | Song H-Q, Pan W, Li R-Q, Yu B, Liu W, Yang M, et al. Rodlike Supramolecular Nanoassemblies of Degradable Poly(Aspartic Acid) Derivatives and Hydroxyl-Rich Polycations for Effective Delivery of Versatile Tumor-Suppressive ncRNAs. Nano Micro Small. 2017;14(9):1703152. [Google Scholar] [CrossRef] |
| 291. | Park KS, Sun X, Aikins ME, Moon JJ. Non-viral COVID-19 vaccine delivery systems. Adv Drug Deliv Rev. 2021;169:137-151. [Google Scholar] [CrossRef] |
| 292. | Landmesser U, Lüscher TF. Advancing RNA-targeted therapy for personalised prevention of coronary disease: focus on ANGPLT3. Eur Heart J. 2020;41(40):3946-3948. [Google Scholar] [CrossRef] |

![]()
Copyright © 2026 Pivot Science Publications Corp. - unless otherwise stated | Terms and Conditions | Privacy Policy




